Фенотип: визначення, приклади, взаємозв'язок з генотипом і генетичну різноманітність
визначення: фенотип - виражені фізичні особливості організму, певні генотипом, випадкової генетичної варіацією і впливом навколишнього середовища.
приклади: такі риси, як колір, висота, розмір, форма і поведінку.
Взаємозв'язок фенотипу і генотипу
Генотип організму визначає його фенотип. Всі живі організми мають ДНК, яка дає інструкції для виробництва молекул, тканин і органів. ДНК містить генетичний код, який також відповідає за напрямок всіх клітинних функцій, включаючи, реплікацію ДНК, синтез білка і перенесення молекул.
Фенотип організму (фізичні риси і поведінку) визначаються їх успадкованими генами. являють собою певні ділянки ДНК, які кодують структуру білків і визначають різні ознаки. Кожен ген розташований на і може існувати в більш ніж одній формі. Ці різні форми називаються алелями, які розташовуються в певних місцях на певних хромосомах. Аллели передаються від батьків до потомства через.
Диплоїдні організми успадковують два аллеля для кожного гена; один аллель від кожного з батьків. Взаємодія між алелями визначають фенотип організму. Якщо організм успадковує два однакових алелі для певної ознаки, він гомозиготний за цією ознакою. гомозиготні особини висловлюють один фенотип для даної ознаки. Якщо організм успадковує два різних аллеля для певної ознаки, він є гетерозиготних за цією ознакою. Гетерозиготні особини можуть висловлювати більш одного фенотипу для даної ознаки.
Риси можуть бути або рецесивними. У схемах успадкування повного домінування фенотип домінуючою риси повністю маскує фенотип рецесивного ознаки. Є також випадки, коли відносини між різними алелями не виявляють повного домінування. При неповному домінуванні домінуюча аллель повністю не маскує іншу аллель. Це призводить до фенотипу, який являє собою суміш фенотипів, які спостерігаються в обох аллелях. При кодоминировании обидва алелі повністю виражені. Це призводить до фенотипу, в якому обидві ознаки спостерігаються незалежно один від одного.
Повний, неповний і кодоминирование
Фенотип і генетичну різноманітність
Може впливати на фенотип. Воно описує зміни генів організмів у популяції. Ці зміни можуть бути результатом мутацій ДНК. Мутації є змінами послідовностей генів в ДНК.
Будь-яка зміна послідовності генів може змінити фенотип, виражений в успадкованих аллелях. Потік генів також сприяє генетичною різноманітністю. Коли нові організми потрапляють в популяцію, вводяться нові гени. Введення нових алелей в генофонд робить можливими нові комбінації генів і різні фенотипи.
Під час утворюються різні комбінації генів. У мейозі випадковим чином поділяються на різні. Передача гена може відбуватися між гомологічними хромосомами через процес перетину. Ця рекомбінація генів може створювати нові фенотип в популяції.
Ми продовжимо обговорювати якісні ознаки, і говорити про зв'язок генотипу і фенотипу, про варіанти прояви менделевских закономірностей, і що за цим стоїть з точки зору наших уявлень про роботу генетичного матеріалу.
Навіщо треба вивчати результати Менделя і Моргана? Перш за все, нам цікаво дізнатися щось про нас самих. Нам хочеться бути багатими, здоровими і щасливими (останні дві ознаки багато в чому генетично контрольовані. Є люди, які відчувають себе зазвичай щасливими, а є такі, які частіше відчувають себе нещасними - ці відчуття пов'язані з певними генетичними характеристиками).
Якщо ми знаємо характер генетичного контролю ознаки, то ми можемо передбачити з певною ймовірністю фенотип на основі генотипу (якщо він відомий). Якщо ми не знаємо, як ознака контролюється, то ми будемо в повному невіданні, і не зможемо нічого сказати щодо ознак майбутніх поколінь. Якщо ми знаємо зв'язок між генотипом і фенотипом, то ми можемо зробити певні передбачення про розвиток ознаки (наприклад, хвороби) і, в деяких випадках, вжити заходів корисні індивіду. Для цього нам треба встановити генотип. Зараз це завдання технічно можна вирішити (оскільки секвенування генома людини проведено), хоча і непомірно дорога.
В реальності нам дано лише спостерігати прояв ознаки в поколіннях і на основі цього створювати модель генетичного контролю формування ознаки, яка може бути вірна лише в даних конкретних умовах. Але, тим не менше, якщо ми створили таку модель, то ми можемо в цьому випадку мати кошти для регуляції якоїсь ознаки, зокрема, вплинути виникнення або перебіг будь-якої хвороби. Таким чином, генетичний контроль і його вивчення має велике практичне, зокрема, медичне, значення. І в основі всього лежать Менделя закономірності, які можуть проявляються по-різному в залежності від конкретних особливостей генотипу і середовища.
Ми зараз розглянемо, які можуть бути умови генотипу і середовища, при яких ці закономірності будуть виглядати не такими, як їх спостерігав Мендель, і чому це відбувається. Мендель спостерігав, що при об'єднанні задатків двох ознак в одному організмі може бути прояв тільки однієї ознаки. Другий задаток не проявляється. Такий тип домінування називається повним.
Перевідкриття законів Менделя призвело до виявлення інших типів домінування. наприклад, неповного домінування, Коли фенотип гетерозиготи є проміжним між двома гомозиготами. Є ще один тип домінування, дуже популярний останнім часом в суді, - кодоминирование - в гетерозиготі проявляються фенотип кожної з гетерозигот. Це явище має місце, зокрема, і у людини. Якщо у вас є татова хромосома і мамина хромосома (а це, безсумнівно, так), і вони різняться в мільйон позицій, що можна виявити різними методами, то це все випадки Кодомінування.
Первинним фенотипом організму є послідовність нуклеотидів молекул його ДНК. На цьому фенотипе будуються всі фенотипи наступних рівнів. Тобто при дослідженні вашої ДНК виявляються все і татові, і мамині ознаки; кожна з молекул ДНК проявляє свою ознаку незалежно від присутності іншої молекули ДНК з іншою ознакою: при секвенування або при розщепленні ДНК будь-якими ферментами видно обидва стану ДНК. Кодомінантність ознаки (маркери самої молекули ДНК) характеризують відмінність між хромосомами і використовуються для ідентифікації особи або встановлення батьківства (число таких випадків дозволених в суді становить кілька сотень на рік).
Коли ми говоримо про генотип і фенотип - це такі крайні єдиного процесу реалізації спадкової інформації в індивідуальному розвитку. Наприклад, гладка або зморшкувата форма горошини, це її фенотип. А генотип - це та специфічна послідовність нуклеотидів, яка в даних умовах визначає, що горошина буде гладкою або зморшкуватою. У 1999 році з менделевскими лініями гороху була проведена наступна робота. Ділянки хромосоми, що відповідають за форму горошини, були клоновані, секвенувати, і було встановлено їх особливості - відмінність послідовностей нуклеотидів - які і визначали розвиток гладкою або зморшкуватою форми горошини.
Зверніть увагу, що форма горошини є кінцевим ознакою, а формування ознаки цього рівня передує прояв ознаки на багатьох попередніх рівнях. По-перше, це наявність (аллель 1) або відсутність (аллель 2) олігосахариду, яке і призводить до тієї або іншій формі горошини. Ще більш глибоким рівнем прояви фенотипу є те, що є відповідний білок (аллель 1), який необхідний для синтезу олігосахариду або це той же білок, але альтернативної структури (аллель 2), при якій олігосахарид не утворюється. Ще більш глибоким ознакою є РНК, з якою синтезується цей білок. Ці РНК різні за послідовності нуклеотидів (аллели 1 і 2), що і робить різними відповідні білки. А ці РНК різні, тому що транскрибується з різних молекул ДНК, татової і маминої, у яких послідовності нуклеотидів в даній позиції різні (аллели 1 і 2). Все це - прояв одного і того ж фенотипу, послідовно реалізується на кожному з рівнів.
Ми маємо право говорити про фенотипе на кожному з цих багатьох рівнів - від конкретних особливостей послідовності нуклеотидів ДНК до форми горошини. При цьому, як тільки ми просуваємося від ДНК вище, тим більше вплив умов навколишнього середовища. Наприклад, можливість функціонування різних алелей на рівні ДНК (транскрипція татової і маминої копії гена) буде мало залежати від температури, а можливість функціонування тих же алелей на рівні білка може критично залежати від температури. При деяких температурах білок (наприклад, аллель 1) буде працювати, а інший (аллель 2) не буде працювати. Як тільки ми висуваємося на більш високий рівень в реалізації фенотипу, з'являється більше можливостей для впливу навколишнього середовища на прояв ознаки.
І навпаки, чим ближче ми просуваємося до генотипу, тим передбачувані зв'язок між генотипом і фенотипом. При ідентифікації людини в судово-медичній експертизі найчастіше використовуються маркери, які розрізняють одні й ті ж ділянки гомологічних (татові і мамині) хромосом. Зазвичай ці відмінності - мікровставкі в даному місці молекули ДНК. Принцип методу такий. Ми можемо розмножити in vitro фрагмент молекули ДНК в потрібному місці хромосоми, наприклад, з координатами від позиції N до позиції N + 100. Цей фрагмент ми вибрали тому, що в ньому у кожної людини в кожній хромосомі є вставка, наприклад, дінуклеотід СА. У кожній хромосомі довжина цієї вставки може бути своя. Припустимо, в одній хромосомі цей дінуклеотід повторений в даному місці 10 разів, а в тому ж місці інший хромосоми - 15 разів. При розмноженні цієї ділянки хромосоми ми отримаємо фрагменти двох довжин 100 + 2 * 10 = 120 пар нуклеотидів і 100 + 2 * 15 = 130 пар нуклеотидів. Фрагменти кожного з цих розмірів відрізняються після їх фракціонування електрофорезом в гелі.
Нехай і тато і мама гетерозіготни з цього локусу, подібно гібридам першого покоління менделевским моногибридного схрещування. Позначимо їх генотипи по характеристичній довжині розмножуються фрагментів ДНК - 120/130. Тоді їхні діти матимуть такі генотипи 1 120/120 (гомозиготи по аллелю 120): 2 120/130 (гетерозиготи): 1 130/130 (гомозиготи по аллелю 130). Природно, присутність в зразку фрагмента ДНК однієї довжини не впливає на можливість детекції фрагмента ДНК іншої довжини, тобто, використовувані маркери ко-домінантні. Вони обидві проявляються. Зрозуміло, що тут проявляються ті ж Менделя закономірності передачі і розщеплення ознак, хоча в якості ознаки ми використовували морфологію самої молекули ДНК - різну довжину фрагмента молекули в даному місці хромосоми.
Після перевідкриття законів Менделя з'ясовувалося, що Мендель то в одному як би як би не правий, то в іншому як би не правий. Виявлялися додаткові обставини, які модифікували прояв менделевских закономірностей.
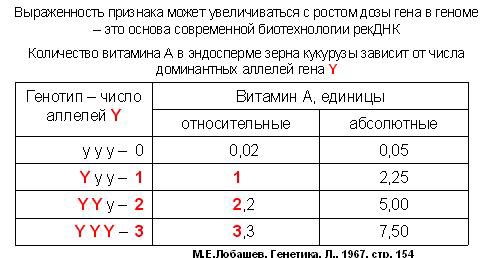
Наприклад, число класів генотипів і фенотипів залежить від плоїдності, навіть якщо алелей всього два. Наприклад, ендосперм в зерні кукурудзи тріплоідний. У нього два генома маминих, а один татів (з пилкового зерна). Отримували рослини кукурудзи, які розрізнялися за кількістю домінантного алеля гена Y. Цей аллель необхідний для утворення вітаміну А. Коли в ендоспермі була одна копія алелі Y, то Якщо відносна кількість вітаміну А при одній копії алелі Y становило одиницю, то при двох копіях - 2 , 2, а при трьох копіях - 3,3. Тобто вираженість конкретного ознаки може залежати від дози гена. А може і не залежати - як у випадку повного домінування одного алеля над іншим. Все залежить від конкретного механізму формування ознаки.
Те, що від дози гена залежить кількість його продукту, використовується в біотехнологіях, як основа всіх сучасних процесів, заснованих на рекомбінантних ДНК. У всіх цих випадках в клітці збільшують дозу гена. Збільшення дози гена призводить до потрібних нам результатами, але клітина від цього може померти. Зазвичай при виробництві це байдуже. Критерієм ефективності в біотехнології є отримання цільового речовини з мінімальними витратами. Втім, якби могли отримати корову, яка б дала відразу цистерну молока, а потім би померла, це було б дуже технологічно, але не етично. Однак з бактеріями так і роблять: вирощують бактерії в ідеальних умовах, а потім включають синтез, який стоїть бактеріям життя.
Наприклад, за допомогою технологій рекомбінантних ДНК сконструйовані бактерії, здатні перетворювати цукор в амінокислоту треонін. Більше половини вуглецю, поглиненого такими бактеріями перетворюється в єдиний цільовий продукт. Менше половини цукру перетворюється в тисячі інших вуглець-містять сполук клітини, а, крім того, витрачається ще й як джерело енергії, щоб все це синтезувати і просто жити. Треонін накопичується в середовищі до концентрації в 100 грам на літр, що близько до межі його розчинності. Світові обсяги виробництва рекомбінантних продуктів складають десятки тисяч тонн на рік (треонін) і навіть сотні тисяч тонн (лізин).
У менделевским випадку кожна ознака формувався під дією як би одного гена, який на інші ознаки не впливав (адже рослина з гладкими насінням могло бути різної висоти, мати різного кольору насіння і квіти і ін.), Насправді формування кожної ознаки вимагає роботи багатьох генів, і більшість генів впливають більш, ніж на один ознака. Наприклад, відносний вміст кожного з трьох метаболітів в ендоспермі кукурудзи (на слайді) залежить від генотипу по кожному з трьох досліджених генів.

Видно, що на кожну ознаку впливає кожен з трьох генів, і таких генів напевно більше. Для формування кожної ознаки потрібно багато генів. Пояснимо це на прикладі слуху. Щоб людина чула, потрібні багато структур вуха: молоточки, наковаленки, барабанні перетинки, і якщо не буде якогось білка, потрібного, щоб сформувати ці структури, то людина не буде чути. Тому глухота у людини може виникати через дефекти в десятках відомих генів (контролюючих багато елементів слухового апарату, передачу сигналу в нервовій тканині т.д.). При цьому різні генотипи можуть приводити до одного фенотипу (глухота). Зовні однакові фенотипи, обумовлені різними генетичними причинами, називаються фенокопії. Мутації глухоти рідкісні і зазвичай рецесивні. Глухота проявляється зазвичай як моногенний аутосомний дефект зі спадкуванням за менделевскому типу. Відомі випадки, коли чоловік і дружина, обидва глухі, хотіли б, щоб і їх майбутня дитина був глухим, щоб він також належав їх спільноті. Цей приклад показує - поняття норми і хвороби відносно. Воно залежить, в тому числі, і від ставлення індивіда до своєї особливості і від ставлення суспільства до цієї особливості.
Менделя закономірності при взаємодії генів (на прикладі глухоти). Припустимо, що глухота визначається рецесивними мутаціями в 10 різних генах і частота цих мутацій в популяції однакова.
Питання 1. Яка ймовірність того, що у двох глухих батьків буде глухий дитина? Рішення. Кожен з батьків - гомозигота по мутації в одному з генів, потрібних для розвитку слуху. Імовірність того, що другий з батьків несе мутацію в тому ж гені, що і перший становить 1/10. Отже, ймовірність того, що дитина від такого шлюбу буде глухим, становить 10%, а того, що він буде чути - 90%. Запис генотипів буде при цьому наступна: батьки ааВВ х АА bb, Потомство першого покоління Аа Bb.
Питання 2. Яка буде доля тих, що чують нащадків від схрещування індивідів першого покоління? (Щоб не бентежити один одного будемо вважати, що цей досвід проводиться, звичайно, не на людях, а на мишах - основному модельному генетичному об'єкті з ссавців). Рішення. Розщеплення по кожній парі алелей відбувається незалежно один від одного. Тобто схрещування Аа Bb х AaBb можна записати як два незалежних схрещування. За гену А: батьки Аа х Аа. Розщеплення у нащадків за фенотипом складе 3: 1 (3 чують до 1 глухому). При цьому у всіх чують є хоча б один домінантний аллель А. Такий генотип записується в формі А-. Генотип глухих нащадків - аа (Гомозиготи по рецесивним аллели). Тоді генотипи відповідні чують і не чують нащадкам схрещування можна уявити як 3 А- : 1аа.
Таким же чином запис нащадків за генотипом при розщепленні алелей по локусу В можна уявити як 3 В- : 1bb.
Оскільки розщеплення по аллелям одного локусу відбувається незалежно від розщеплення по аллелям іншого локусу, то генотип нащадків при розщепленні за двома локусами одночасно можна записати в наступному вигляді:
(3А- : 1аа) Х (3 В- : 1bb) = 9A-B- : 3aaB- : 3A-bb : 1aabb
Щоб індивід чув, йому необхідно мати хоча б один аллель дикого типу по кожному з локусів. Таких індивідів 9 (генотипи А-В-). Щоб індивід не чув, досить гомозиготности за допомогою одного з двох рецесивних генів. Таких індивідів 7 (генотипи aaB-, A-bb і aabb)
Таким чином, при схрещуванні дигетерозигот співвідношення чують нащадків до нечуючих складе 9: 7.
Окремий ген може впливати на кілька ознак, у розвиток яких він залучений. Такий ефект дії гена називається плейотропних (множинним). Наприклад, альбінізм викликається дефектом одного гена, контролюючого утворення меланіну. При цьому змінюються одночасно такі ознаки як колір волосся (білий), колір шкіри (білий) і колір очей, (він буде корисним від кольору кровоносних судин, що лежать за райдужною оболонкою ока). Безліч ознак визначається геном SRY, який контролює формування всіх ознак чоловічої статі у ссавців. перенесення гена SRY з Y-хромосоми самця в геном самки миші, приводив до розвитку особини з величезною кількістю зовні спостережуваних ознак чоловічої статі.
У Менделя вийшли однакові чисельні співвідношення при розщепленні алелей багатьох пар ознак. Це зокрема мало на увазі однакову виживання індивідів всіх генотипів, але це може бути і не так. Буває, що гомозигота по якомусь ознакою не виживає. Наприклад, жовте забарвлення у мишей може бути обумовлена гетерозиготністю по одній з пар алелей (слайд). При схрещуванні таких гетерозигот один з одним слід було б очікувати розщеплення за цією ознакою співвідношенні 3: 1. Однак, спостерігається розщеплення 2: 1, тобто 2 жовтих до 1 білої (рецесивна гомозигота). Показано, що домінантна (за забарвленням) гомозигота не виживає вже на ембріональній стадії. Цей аллель одночасно є рецесивною леталей (тобто рецессивной мутацією, що приводить до загибелі організму).

На наступному слайді показані типи груп крові, їх успадкованого. Цей приклад наведено для того, щоб сказати, що у одного гена може бути не тільки два алелі. У Менделя їх було всього два для кожної ознаки: наприклад жовта і зелена забарвлення, третього не дано. Але насправді алелей по кожному локусу може бути багато. Групи крові розрізняються по тому, який тип антигену ми виробляємо. Може вироблятися антиген типу А або В, Але якщо одна хромосома відповідальна за тип А, А інша за тип В, То будуть вироблятися обидва типи антигену. Перед нами випадок Кодомінування на рівні білка, коли наявність одного типу антигену не перешкоджає наявності іншого. Четвертий тип - коли антигени взагалі не виробляються. Яке практичне значення це має для нас? Це були перші генетичні дослідження, проведені під час першої світової війни в австрійській армії, так як під час війни завжди була гостра необхідність переливання крові, яке не завжди виявлялося вдалим, тобто людина від переливання крові іноді помирав.

Переливати кров найбезпечніше між людьми, у яких однакова група крові. Антигени груп крові у них однакові, антитіл проти них немає і аглютинація (злипання клітин крові) не відбудеться.
Якщо донором (тобто здають кров) є людина, у якого жодного з антигенів немає (група крові 0 ), То його кров можна переливати будь-кому - вона не буде викликати аглютинації у реципієнтів (тих, кому кров переливають) з будь-яким типом крові. Таким чином, люди з типом крові 0 є універсальними донорами. І навпаки, в крові людей з групою АВ не містяться антитіла ні на один з цих антигенів (ні на А, Ні на В) І ці люди щасливі бути універсальними реципієнтами - переливати кров їм можна від людей своєї та всіх інших груп крові. Тобто людина з групою крові АВ може прийняти кров з будь-яким типом. Найгірший випадок для реципієнта - нульова група крові, яка буде реагувати з групами крові А, В і АВ.
Два різних гена можуть перебувати в різних локусах і впливати на один і той самий ознака. Наприклад, у дрозофіли можуть бути різний колір очей, від коричневого (рецесивна гомозигота по одному гену), через різні відтінки червоного до білого (рецесивна гомозигота по іншому гену). Червоні очі має муха дикого типу (наприклад, гомозигота за всіма аллелям дикого типу), а також гетерозигота за двома генами, кожен з яких потрібен для пігментації ока (див. Рис. Зліва). Взаємодія двох генів в такому випадку називають комплементарностью (додатково). При схрещуванні дигетерозигот дає розщеплення за фенотипом у відношенні 9: 3: 3: 1. З чотирьох фенотипічних класів за забарвленням очі найчисленніший - 9, це забарвлення дикого типу, а самий нечисленний - 1, відсутність забарвлення з дефект обох генів, що визначають утворення різних пігментів.

Така ж ситуація у людини з забарвленням волосся, у нас є різні типи меланіну: ЕУ і фікомеланін, які в різному поєднанні дають різні відтінки, наприклад рудий.
Справа на малюнку наведено той же приклад, тільки на рослинах . Гени в різних локусах можуть давати внесок в розвиток однієї і тієї ж фенотипической характеристики: у перцю 2 різних гена потрібні для утворення двох різних пігментів, що призводить до розщеплення по типу дигибридного схрещування. Це використовується при отриманні певного кольору перцю. Щоб отримає потомство з потрібним кольором плода необхідно знати генотипи батьків і правильно їх підбирати для схрещування.

На тварин це застосовується на хутрових звірів в звероводческих фермах. За рахунок певного добору алелей десятків відомих генів, що визначають забарвлення шерсті, можна отримати велику різноманітність забарвлень шкур хутрових звірів, причому заздалегідь планувати скільки і яких типів забарвлення вийде.
Якщо два різних гена взаємодіють при формуванні ознаки
і їх дія складається, то така дія генів називають кумулятивним (накопичується). Наприклад, ступінь червоного забарвлення зерна пшениці визначається двома різними генами, в кожному з яких є неповністю домінантний аллель А.
Таким чином, спостерігається багато варіантів чисельних співвідношень класів в потомстві дигибридного схрещування в залежності від типу взаємодії генів. Якщо взаємодії немає, то ставлення буде 9: 3: 3: 1. Може бути так, що поява в генотипі рецесивного гена а веде до того, що ген В в фенотипі не проявляється. Тоді співвідношення буде 9: 3: 4. Те ж, але з домінантним геном - буде 12: 3: 1. І т.д. Тобто це показує, що Мендель був прав тільки в досить окремому випадку, коли гени не впливають один на одного, але може бути ще багато інших варіантів.

Породи собак: для того, щоб отримати якісь певні зафіксовані ознаки породи, проводили близькоспоріднені схрещування, поки ці ознаки що закріплювалися в гомозиготному стані, і далі при схрещуванні собак однієї породи розщеплення за цією ознакою не може відбуватися - ознака стає стабільним, типовим для породи.

Кожна порода собак має свої ознаками, і ці ознаки стабільні в поколіннях, оскільки кожну породу характеризує свій набір гомозиготних локусів, що визначають ці ознаки. У кожної породи собак як свій набір гомозиготних локусів (у інших порід вони можуть бути і гетерозиготних). Однак у одних порід даний локус гомозиготен по одним аллелям, а інших той же локус може бути гомозиготен по іншим аллелям, що і визначає стабільні відмінності між породами (наприклад, по окрасу). Ясно, що у кожної породи якісь локуси залишаються гетерозиготними, і собаки однієї породи за цими ознаками будуть відрізнятися, але це ті ознаки, які дану породу не визначають.

Якщо схрестити собак різних порід, то вийде дворняжка. У неї за більшістю генів гетерозиготное стан, відповідне предковим формам. Дарвін свого часу підмітив, що якщо схрещувати різні породи голубів, виходить весь час сизий. Це відбувається через втрату у гібрида гомозиготности характерною для кожної з батьківських порід. А в гетерозиготних локусах один з алелів, як правило, виявляється дикого типу, тому й виглядають гібриди Сизара - це дикий тип для всіх порід голубів.
За генами забарвлення хутра у норки в даний час встановлено близько 20 серій множинних алелей. При схрещуванні гетерозиготних норок між собою в другому поколінні буде розщеплення, типове для дигибридного схрещування в співвідношенні 9: 3: 3: 1, або 56% норок з дикої забарвленням хутра, 19% - алеутських, 19% - платинових і 6% з сапфіровою забарвленням хутра. Вавилов сформулював закон гомологічних рядів: якщо у якогось виду є якийсь набір варіантів ознак, і ми розглядаємо близькоспоріднений вид, то у нього теж може бути виявлений той же набір ознак. Цей закон заснований на тому, що у близькоспоріднених видів і набір алелей в ДНК може бути схожий.

Гени можуть перебувати не тільки в ядрі. Гени можуть успадковуватися однородітельскі, наприклад, по материнській лінії: забарвлення листа визначається генами хлоропластів, а не ядра, і успадковується тільки від матері. Тобто, яка б не була чоловіча особина, потомство буде все однотипно - все в матір і тільки в неї.
У людини теж є ознаки, які визначаються мітохондріями, тобто успадковуються по материнській лінії. Мітохондрії знаходяться в цитоплазмі клітини, яка дістається нащадкам тільки від матері. До мутацій і хвороб пов'язаних з мітохондріями відносяться, наприклад, дефекти зору і дефекти енергетичного обміну.
Частина ядерних генів також успадковується тільки однородітельскі. Це гени Y-хромосоми, яка успадковується тільки від батька до сина. Ті гени в Y-хромосомі, які не мають пари в Х-хромосомі, успадковуються однородітельскі - від батька до сина. У людини один з таких генів визначає ознака «волохаті вуха» - у жінок така ознака не проявляється - гена цього немає, немає і його алелі, через якого вуха волохаті.
Прояв ознаки може залежати також від статі індивіда. Існують ознаки, гени яких можуть знаходитися в аутосомах або статевих хромосомах обох статей, але проявляються лише у одного з них. Такі ознаки називають обмеженими підлогою. Наприклад, бики несуть гени, що визначають молочність дочок, але гени свою дію у биків не виявляють. Гени казеїну - білків молока - це випадок найбільш суворого контролю транскрипції обмеженою підлогою. Вони діють у особин як жіночого, так і чоловічої статі. Різниця рівнів експресії між ними становить 1 на 100 000. Півні також несуть гени несучості і розміру яєць дочок, хоча у птахів дію цих генів придушене. Коли проводиться селекція, враховується цей факт. Тут істотний генетичний контроль ознак.
Як прояв дії гена, так і тип домінування може залежати від статі. Плішивість у людини успадковується аутосомно і моногенно. Аллель плешивости у чоловіків домінує, а у жінок рецессивен. У чоловіків для облисіння досить присутності одного аллеля плешивости. При цьому частота аллеля дорівнює частоті фенотипу. У жінок для облисіння необхідна присутність обох алелей плешивости.

Дайте відповідь на кілька цікавих запитань.
Скільки жінок будуть лисими, якщо у чоловіків лисий кожен десятий? Відповідь. Частота фенотипу плешивости у жінок дорівнює квадрату частоти алелі, отже, одна зі ста.
Чи може у неплешівих батьків бути плішива дочка? Відповідь. Ні: так як батько неплешівий, значить у нього немає такого алеля (у чоловіка аллель плешивости домінантний і мав проявитися) - обидва алелі визначають нормальну обволосненность. Значить, дочка обов'язково отримає аллель волосатості від батька, а цей аллель у жінок (на відміну від чоловіків) домінантою. Лисою вона не буде незалежно від того, який аллель прийде від матері.
Чи може у неплешівих батьків бути лисий син? Відповідь. Так: якщо у мами є аллель плешивости він рецессивен і не проявиться, він може передатися синові у якого цей аллель проявиться як домінантний.
Якщо у неплешівих батьків один син лисий, наскільки ймовірним є те, що інший теж буде неплешівий? відповідь: 50%. Аллель плешивости не у батька - інакше б у нього цей аллель проявився як домінантний. Значить аллель плешивости у матері. Раз вона волохата, то другий аллель у неї - нормальна обволосненность, а аллель плешивости не проявляється, так як у жінок він рецессивен. Отже, мати - гетерозигота - 1 аллель і неї - плішивість, 1 - норма. Тому розщеплення за цією ознакою у її синів буде 1: 1 (50% плішивих)
Так генетика вирішує практично важливі для нас завдання.
Експресія деяких генотипів може залежати від зовнішніх умов. Нижче показані два кролика, один з яких з темними плямами. Аллель гімалайської забарвлення у кролика температурочувствітелен. При підвищеній температурі білок не функціональний і необхідний пігмент не утворюється, а при нормальній температурі виходить кролик, у якого деякі ділянки шкіри пофарбовані.

Мутанти, прояв яких залежить від умов середовища, зіграли дуже важливу роль в біології взагалі і в молекулярній біології зокрема, в розумінні того, як працює генетичний матеріал. Багато генетичні феномени, були відкриті і досліджені після отримання та використання умовно летальних мутантів (умовно смертельні). Найбільш поширені мутанти такого типу - температурочувствітельной. Отримано мутанти багатьох мікроорганізмів і вірусів, які не здатні жити при підвищеній температурі ( ts-мутанта). У такого мутанта структура білка при підвищеній температурі змінена, і білок не функціональний (леталь), а при нормальній температурі структура звичайна, і білок функціональний. При цьому можливо досліджувати особливості функціонування білка і його ролі в формуванні будь-якої структури або складної функції. Для цього клітини ростять до потрібної стадії, в умовах, коли білок функціональний, потім розділяють клітини (зазвичай суспензію) на дві частини, в одній температура залишається нормальною, а в інший -піднімають, так що білок втрачає функціональність.
Є також інший тип умовно летальних мутацій - нонсенс-мутації, летальні на одному генетичному тлі і нелетальної на іншому. Нонсенс-мутація призводить до зміни кодону для будь-якої амінокислоти в даному білку на стоп-кодон трансляції. Білок при цьому утворюється укорочений і нефункціональний. Нонсенс-мутація пригнічується (супрессірует), в тих клітинах, де утворюється супрессорная тРНК. Супресорна тРНК, це змінена тРНК, у якій антикодон як і раніше дізнається стоп-кодон в матричної РНК, але замість термінації (закінчення) трансляції білка на цій ділянці відбувається підстановка певного амінокислотного залишку. Білок з нонсенс-мутацією синтезується повнорозмірним і функціональним в присутності тРНК.
При створенні моделі успадкування ознаки для з'ясування його генетичного контролю починають з перевірки типу успадкування за базовою, найпростішою схемою - менделевской і, якщо відповідності немає, то додають до цієї схеми додаткові умови, зрозумілі на сьогодні:
- нерівну ймовірність утворення і поєднання гамет і виживання потомства;
- однородітельскій тип спадкування (Y - волохаті вуха, мтДНК - хвороба Лебера)
- взаємодія алелів: наявність домінування;
- взаємодія генів;
- зчеплення генів;
- обмеження ознаки підлогою;
- ймовірність прояву ознаки при даному генотипі;
- вплив середовища (не завжди формалізованих).
Нагадаємо, що менделевским характер успадкування відноситься до ознак якісним. Скільки Меделірующіх ознак відомо у людини? Вони зафіксовані в базі даних OMIM ( «Менделівська успадкування у людини»). Їх там перераховано близько п'яти тисяч ознак. А ми у себе можемо нарахувати набагато більше ознак. Величезна кількість інших наших ознак залежить від дії багатьох генів. Тут слід зазначити важливий ідеологічний аспект. Загальної моделі, за якої відбувається формування ознак під впливом багатьох генів, зараз немає. Напевно, її і бути не може. Щоб зрозуміти, як даний фенотип виникає на тлі даного генотипу, нам потрібно ще дізнатися, як саме реалізується кожен етап передачі генетичної інформації: що відбувається з модифікацією ДНК, з якою з багатьох можливих варіантів РНК утворюється з даної ділянки генетичної інформації, як модифікується білок, утворений з цієї мРНК. У ДНК закладена лише можливість усіх цих варіантів реалізації генетичної інформації. Чи відбудеться це чи ні, залежить від середовища. Поняття середовища ми поки толком формалізувати не можемо. Але якщо, подібно структурі ДНК, ми їх будемо знати структуру і модифікацію РНК і білків, то розуміння зв'язку генотипу з фенотипом буде набагато глибшим, а предсказательная сила наших знань буде більше. Як би там не було, одного знання генотипу часто недостатньо для передбачення фенотипу. Тому ми повинні вивчати наступні етапи експресії генетичної інформації.
У заключній частині лекції йтиметься про визначення статі і ознаках, зчеплених зі статтю. Нижче показано, як виглядають статеві хромосоми людини. Х-хромосома по довжині ДНК більше ніж Y-хромосома приблизно в три рази.

А так виглядають клітини, що транспортують ці хромосоми. Маленька утворена організмом генотипу XY, А велика - генотипу ХХ. Таке співвідношення фізичних розмірів мають статеві клітини.
Так виглядає людина після того, як він вже почав розвиватися:

Це голка, і на її кінці знаходиться кілька поділилися клітин, отриманих із заплідненої яйцеклітини. Такий наш розмір на стадії 4-5-ого поділу.
Ще раз нагадаю, що у нас цикл розвитку двохстадійний. На нижній частині малюнка зображена диплоидная стадія, період нашої довгого і щасливого життя, яка з точки зору генетичного процесу закінчується мейозом, після якого ми починаємо жити як би в наступній стадії, гаплоидной. Це гамети, у нас вони не мають здатність до самостійного існування, хоча у багатьох рослин саме гаплоїдії є основною частиною життєвого циклу. Існування гамет закінчується заплідненням і появою знову диплоидного організму.

Х і Y хромосоми сильно відрізняються. Вони гомологични тільки в так званих псевдоаутосомних зонах, які присутні і в батькових і в маминих хромосомах, як і звичайні аутосоми. Спермії можуть бути двох типів: Х і Y, а яйцеклітини тільки одного типу - Х і Х. Тому розщеплення виходить 1: 1 (дві жінки і два чоловіки). Саме розщеплення 1: 1 по підлозі на першому етапі дослідження ознаки, зчепленого з підлогою, - забарвлення очей у дрозофіли - призвело Моргана до думки про те, що гени знаходяться в хромосомах. Пол визначається хромосомами і гени знаходяться саме в них.

Не у всіх організмів, на відміну від людини, стать однозначно визначається при злитті гамет. Нижче показаний молюск, у якого в нижній частині жіночі клітини, а у верхній частині тільки чоловічі. У деяких плазунів чоловіча стать не формується при підвищеній температурі (зокрема, у крокодилів), утворюються тільки жіночі організми.

У дрозофіли всього чотири пари хромосом, три пари хромосом називаються аутосоми і одна пара статевих хромосом. У самок набір хромосом ХХ, А у самця Х Y - як і у людини (у самки на кінці черевця знаходиться яйцеклад, тому у неї черевце загострене, а у самця воно овальне, - так їх відрізняють).

У плодової мушки підлогу визначається не стільки наявністю тієї чи іншої статевої хромосоми, а ставленням числа Х-хромосом до числа гаплоїднийнаборів аутосом (А). Це видно на схемі нижче. набір ХХ+2А (2 Х-хромосоми і 2 набору аутосом) дає співвідношення 1,0 - таке співвідношення є нормою для самок. набір Х Y+2А (1 Х-хромосома і 2 набору аутосом) дає співвідношення 0,5 - таке співвідношення є нормою для чоловічої статі. Якщо виявляється, що Y-хромосома загублена (набір 1 Х+2А, Співвідношення 0,5), то утворюється чоловічий організм, хоч Y-хромосоми і немає. А якщо на тлі двох Х-хромосом додалася Y-хромосома (набір 2 Х+2А), Стать формується жіночий, хоч Y-хромосома і присутній. У людини, скільки б не було Х-хромосом, якщо є хоч одна Y-хромосома, то фенотип буде чоловічий.
Таким чином, у людини і мухи, при зовні однаковою хромосомної формулою норми (гомогаметний підлогу, ХХ, Жіночий; гетерогаметний підлогу, Х Y - чоловічий), механізм генетичного контролю освіти статі різний.
У птахів і ряду інших груп організмів гомогаметний підлогу ( ХХ) - чоловічий, а гетерогамтний підлогу ( XY) - жіночий, що протилежно тому типу хромосомного визначення статі, який виявився зафіксований еволюцією в лінії людини і інших ссавців.

На наступному слайді показано розташування SRY-гена, що відповідає за формування ознак чоловічої статі. Уже говорилося, що якщо в геном миші, яка повинна розвинутися самкою, пересадити SRY-ген, то у такого організму розвинеться чоловічий фенотип. Людина майже не відрізняється від миші в цьому плані. У людини відомі випадки, коли індивід каріотіпіческі (тобто за типом хромосом в ядрі) відповідав жінці (зокрема з двома ХХ хромосомами і без спостерігається Y-хромосоми), але виявляв фенотип чоловіки. У таких індивідів була виявлена хромосомна перебудова - ген SRY був транслоцироваться (переміщений) на одну з аутосом.

На внизу на малюнку представлений випадок відхилення за складом статевих хромосом - синдром Тернера: індивід генотипу Х0 (Y-хромосоми немає, а Х-хромосома одна). Фенотип індивіда - жіночий. Відхилень у поведінці немає, хоча при більшому дисбалансі в генотипі спостерігаються проблеми в розумовому розвитку.
Інший випадок відхилення за складом статевих хромосом - це синдром Клайнфельтера: генотип XXY, Фенотип чоловічої. Хоча дві Х-хромосоми присутні, як в нормі у жінок. Тобто для визначення статі у людини важливо, щоб була присутня Y-хромосома.

Нагадаємо, що гени, локалізовані в Y-хромосомі і не мають пари в X-хромосомі, успадковуються тільки від батька до сина. У людини аллель одного з таких генів визначає ознака «волохаті вуха». Через мати до дитини можуть передаватися гени, що знаходяться в мітохондріях, але тому що генів в мітохондріях на три порядки менше ніж в ядрі, то і відхилень, що передаються тільки материнській лінії лише тисячна частина від усіх відомих для людини.
Надлишок або нестача якийсь хромосоми в геномі призводить до дуже серйозних порушень у людини. Для прикладу можна навести трисомії по 21-ій хромосомі - синдром Дауна. Причина захворювань при втраті або надлишку будь-хромосоми - дисбаланс в кількості продуктів генів цієї хромосоми щодо кількості продуктів генів інших хромосом (всі - по дві). Дисбаланс по всіх великих хромосомами людини взагалі не сумісний з життям.
Х хромосома одна з великих хромосом людини. У жінок таких хромосом дві, а у чоловіків - одна. Але обидві статі живі і здорові - хоч на себе подивись. Чому різниця в числі копій Х-хромосоми не смертельна, на відміну від ситуації з іншими хромосомами?
Щоб збалансувати дію генів Х хромосоми у чоловіків (де Х-хромосома одна) і у жінок (де Х-хромосоми дві) природа передбачила інактивацію однієї з Х-хромосом в кожній клітині жіночого організму. У жінок одна Х-хромосома інактивована, тобто гени в ній не працюють, а працюють тільки в іншій хромосомі - НЕ інактивованої. Інактівіація викликана компактизації хромосоми навіть в інтерфазі. Тому компактізірованная хромосома видно при фарбуванні - у вигляді так званого тільця Барра (слайд). Це один з діагностичних ознак жіночої статі у ссавців, що у людини, що у слона. Некомпактізованная Х-хромосома, як і всі аутосоми, не фарбується й не видно в інтерфазі.
На наступному малюнку показана кішка черепаховій забарвлення. Оскільки це кішка, а не кіт, то генотип у неї ХХ. У кожній клітині жіночого організму одна з Х-хромосом інактивована (її видно як тільце Барра). На Х-хромосомі у кішки знаходиться один з генів, що визначають забарвлення шерсті. У цього гена відомі різні аллели, тобто визначають різний колір шерсті. Кішка на малюнку гетерозиготна за цими аллелям. Одна з Х-хромосом в кожній клітині інактивована, тому тільки один з двох кольорів може проявиться в волосині, сформованому з даної клітини. Тому одні волоски одного кольору, а інші - іншого. Яка з Х-хромосом інактивується (батькова або мамина), і, отже, аллель якого кольору буде працювати - справа випадку. Тому така кішка і має черепахову забарвлення. До речі, сини у неї будуть двох різних кольорів в співвідношенні 1: 1.

Ще один доказ того, що гени знаходяться в хромосомах, було отримано на мухах дрозофилах, званих латеральними (бічними, «сторонніми») гінандроморфамі. У таких мух після першого поділу яйцеклітини в одній з клітин була втрачена Х-хромосома. Ця клітина при наступних розподілах дала клітини з генотипом Х0, Тому половина тіла таких мух сформована як у самця і у її всіх клітинах (на малюнку - права половина) цитологічних спостерігається втрата однієї з Х-хромосом. Ліва половина тіла, яка відбувається з клітин з нормальним генотипом ХХ, Формується як самка.

У представленому на малюнку випадку муха була гетерозиготна по гену забарвлення очі і по гену, що впливає на форму крила. Ці гени розташовані в Х-хромосомі. Тому праве око мухи непофарбована: проявилася рецесивна мутація white, Присутня в одній з Х-хромосом. Крило змінено за формою. Ліве око забарвлений і крило нормальної форми, оскільки в ньому залишилися обидві Х-хромосоми, в одній з яких присутній рецесивний ген white, А в іншій - домінантний ген w +.
При аналізі за фенотипом виписують можливі варіанти генотипів нащадків з різними поєднаннями ознак кольору шерсті з довжиною вовни. Потім для кожного поєднання ознак записують відповідні йому генотипи. Тому, визначивши фенотипічніпрояв кожного з вище записаних генотипів, згрупуємо всі їх за належністю до якогось певного поєднання ознак:
|
генотипи | ||
|
нерудих, веснянкуватий нерудих, норма рудий, веснянкуватий рудий, норма |
1ААВВ, 2ААВв, 2АаВВ, 4АаВв 1ААвв, 2Аавв 1ааВВ, 2ааВв |
Частка нерудих веснянкуватих дітей буде 9 або 56,25%.
9. Загальні формули розщеплення
Якщо гени, що відповідають за прояв двох НЕ альтернативних ознак, знаходяться в негомологічних хромосомах, то при мейозі вони потраплять в гамети незалежно один від одного. Тому при схрещуванні двох дигетерозигот розщеплення за генотипом:
1Аавв: 2ааВв: 1аАвв: 2АаВв: 4АаВв: 2Аавв: 1ааВв: 2ааВв: 1аавв
є результатом двох незалежних розщеплення
1АА: 2Аа: 1аа і 1ВВ: 2ВВ: 1вв
Математично його можна виразити у вигляді твору:
(1АА: 2Аа: 1аа) і (1ВВ: 2ВВ: 1вв)
(1: 2: 1) 2
Такий запис показує, що серед особин з генотипом АА одна частина несе гени ВВ, дві частини гени Вв, одна частина - гени ст. Аналогічні співвідношення особин по набору генів В будуть для генотипів Аа і аа.
Якщо особини аналізуються за кількома ознаками, то загальна формула розщеплення при схрещуванні повністю гетерозиготних особин буде (1 : 2: 1) n, де n - число пар альтернативних ознак. Максимальне значення n дорівнює числу пар гомологічних хромосом.
Аналогічно іде справа з розщепленням за фенотипом. В основі також лежить моногибридное розщеплення 3 : 1. При схрещуванні двох дігетерозіготних організмів для незалежних розщеплення 3 : 1 дають розщеплення 9 : 3: 3: 1. Загальна формула розщеплення за фенотипом буде (3 : 1).
Знаючи формули розщеплення, годі й составлять грати Пеннета.
Приклад 9.1. Деякі форми катаракти і глухонімоти у людини передаються як аутосомних рецесивних ознаки. Яка ймовірність народження дитини з двома аномаліями в родині, де обоє батьків гетерозиготні по обидва парам генів.
Рішення:
ознака: ген
без катаракти : А Р АаВв х АаВв
катаракта : а здоровий здорова
глухонімоти : В
глухонімота : в F 1 аавв -?
Дитина з двома аномаліями має генотип - аавв. Відомо, що при схрещуванні двох дигетерозигот серед особин першого покоління спостерігається розщеплення за фенотипом. Формула розщеплення виглядає як 9: 3: 3: 1. З цієї формули видно, що на частку дігомозіготного по рецесивним генам організму доводиться 1 частина. Це означає, що ймовірність появи дитини з двома аномаліями дорівнює 1/16 або 6,25%.
Можна підійти до вирішення завдань на ді-і полігібридне схрещування ще простіше. Для цього необхідно знати лише ймовірності прояви тих чи інших генотипів або фенотипів при різних варіантах моногибридного схрещування.
Так, при моногібрідномсхрещуванні гетерозигот (Аа) ймовірність появи потомства з генотипом АА дорівнює ¼, з генотипом Аа - ½, з генотипом аа I / 4, а ймовірність появи особин з ознаками домінантного гена дорівнює ¾, з ознакою рецесивного гена - ¼.
При схрещуванні дигетерозигот (АаВв) ймовірність появи особин з генотипом аа дорівнює - ¼, з генотипом ст - теж ¼.
Щоб знайти ймовірність збігу двох незалежних один від одного явищ, потрібно перемножити ймовірності кожного з них між собою. Наприклад, для визначення ймовірності появи особин з двома рецесивними ознаками (аавв) треба ¼ помножити на ¼, що дасть в результаті 1/16.
Приклад 9.2. У людини ген карих очей домінує над блакитними очима, А вміння володіти переважно правою рукою - над ліворукістю. Неалельні гени розташовані в негомологічних хромосомах. Блакитноокий правша одружився на кароокою правші. У них народилися двоє дітей: кароокий лівша і блакитноокий правша. Визначте ймовірність того, що їх наступний дитина буде блакитноокий і буде володіти переважно лівою рукою.
Короткий запис умови задачі матиме такий вигляд:
ознака: ген
карі очі : А Р А В х Аав
блакитні очі : а кароока блакитноокий
праворукість : У праворуких праворукий
ліворукість : в
F 1 А ст Аав
кароокий блакитноокий
леворукий праворукий
Рішення:
Перш ніж приступити до розрахунку ймовірності народження наступної дитини з генотипом - аавв, необхідно визначити генотипи їхніх дітей. Наявність в генотипі дітей двох пар рецесивних генів (аа і ст) свідчить про те, що обоє батьків несуть гени а і в. Таким чином генотипи матері буде АаВв, батька - аавв.
Імовірність народження наступної дитини з певним генотипом не залежить від того, якими ознаками мають вже народилися діти. Тому рішення задачі зводиться до визначення ймовірності народження у цієї пари блакитнооких лівшів.
Р АаВв х аавв
р (аа) = 1/2 р (ст) = 1/4 р (аавв) = 1/2 х ¼ = 1/8
Імовірність появи генотипу аа при схрещуванні особин з генотипами Аа і аа дорівнює ½, ймовірність появи генотипу ст при схрещуванні Вв і Вв - ¼. Тому ймовірність зустрічі цих двох генотипів в одному організмі буде ½ х ¼ = 1/8.
відповідьІмовірність народження в сім'ї дитини з блакитними очима, що володіє переважно лівою рукою, дорівнює 1/8.
Користуватися цим прийомом слід тільки при повному оволодінні навичкою звичайного рішення задач.
Як ми бачили, існують ознаки, які мають просте однозначне відповідність генам (групи крові, наприклад), і середовище не робить ніякого впливу на прояв таких ознак. Людина, що володіє алелями групи крові А, при будь-яких обставинах буде мати цю групу крові, в якому середовищі вона не перебувала. Але такі повністю генетично детерміновані ознаки є скоріше винятком, ніж правилом. Більшість же ознак людини є результат взаємодії генів з факторами середовища. Ознаки, для яких характерна безперервна мінливість, як правило, обумовлені спільною дією багатьох генів і факторів середовища. Наследуемость таких ознак називають полігенною.
У генетиці існують два дуже важливих поняття. Це поняття генотип і фенотип. Ми вже знаємо, що спадкова конституція складається з великої кількості різних генів. Вся сукупність генів даного організму називається його генотипом, тобто поняття генотипу ідентично поняттю генетичної конституції. Свій генотип (набір генів) кожна людина отримує в момент зачаття і несе його без будь-яких змін через все своє життя. Активність генів може змінюватися, але їх склад залишається незмінним.
Від поняття генотип слід відрізняти інше подібне поняття - геном. Геномом називається сукупність генів, характерна для гаплоидного набору хромосом особини даного виду. На відміну від генотипу геном є характеристикою виду, а не окремої особини.
Фенотип ж являє собою будь-які прояви організму в кожен момент його життя. Фенотип включає в себе і зовнішній вигляд, і внутрішню будову, і фізіологічні реакції, і будь-які форми поведінки, які спостерігаються в поточний момент. Наприклад, вже згадувані групи крові системи АВ0 - це приклад фенотипу на фізіолого-біохімічному рівні. Хоча на перший погляд багатьом здається, що група крові - це генотип, оскільки вона чітко визначається дією генів і не залежить від середовища, однак це лише прояв дії генів, і тому має бути віднесено до категорії фенотипів. Згадаймо, що представники груп крові А чи В можуть мати різні генотипи (гомозиготні і гетерозиготні). Складними фенотипами є все поведінкові прояви. Наприклад, почерк, який відрізняє даного індивіда, є його поведінковим проявом і також відноситься до категорії фенотипів. Якщо група крові протягом життя не змінюється, то почерк у міру тренування навичок письма зазнає значних змін.
Якщо генотипи успадковуються і залишаються незмінними протягом життя індивіда, то фенотип здебільшого не успадковуються - вони розвиваються і є наслідком наших генотипів лише до певної міри, оскільки велику роль в становленні фенотипів грають умови зовнішнього середовища.
Весь процес розвитку від заплідненої яйцеклітини до дорослого організму відбувається не тільки під безперервним регулюючим впливом генотипу, але й під впливом безлічі різних умов середовища, в яких знаходиться зростаючий організм. Тому надзвичайна мінливість, властива живим організмам, обумовлена не тільки величезною різноманітністю генотипів, що виникають внаслідок рекомбінації генів і мутаційного процесу, але в значній мірі пояснюється і тим, що окремі індивіди розвиваються в розрізняються умовах середовища.
З давніх-давен йде полеміка про те, що важливіше для формування організму - середовище або генетична конституція. Особливо гострі суперечки розгораються там, де справа стосується поведінки людини, його психологічних характеристик - темпераменту, розумових здібностей, рис особистості. Не випадково, що саме з питання про природу розумової обдарованості почалися дослідження в галузі генетики людини. Ф. Гальтон першим в науковому трактаті поставив поруч два поняття, які в тій чи іншій формі не сходять зі сторінок наукової літератури до наших днів. Це поняття - "nature and nurture", тобто "природа і умови виховання".
Генетиків, і генетиків поведінки зокрема, часто дорікають в запереченні ролі середовища. Однак такий закид абсолютно необгрунтований. Одним з основних постулатів генетики є теза про те, що фенотип являє собою результат взаємодії генотипу і середовища. У процесі цієї взаємодії і виникає те різноманіття фенотипічних проявів, яке характерно для більшості ознак людини, що відносяться до категорії кількісних і утворюють безперервний ряд мінливості.
Генотип- сукупність всіх генів організму, який він отримує від батьків.
Фенотіп- сукупність зовнішніх і внутрішніх ознак організму, які формуються в процесі взаємодії генотипу і факторів навколишнього середовища.
Каріотіп- сукупність диплоїдного набору хромосом соматичних клітин певного біологічного виду, який характеризується постійністю числа, форми, розміру.
СПІВВІДНОШЕННЯ Генотип і фенотип. Сукупність усіх генів організму називається генотипом. Генотип є взаємодіють один з одним і впливають один на одного сукупності генів. Кожен ген відчуває на собі вплив інших генів генотипу і сам робить на них вплив, тому один і той же ген в різних генотипах може виявлятися по-різному.
Сукупність усіх властивостей і ознак організму називається фенотипом. Фенотип розвивається на базі певного генотипу в результаті взаємодії з умовами зовнішнього середовища. Організми, що мають однаковий генотип, можуть відрізнятися один від одного в залежності від умов розвитку та існування. Окремий ознака називається феном. До фенотипическим ознаками відносяться не тільки зовнішні ознаки (колір очей, волосся, форма носа, забарвлення квіток тощо), а й анатомічні (обсяг шлунка, будова печінки тощо), біохімічні (концентрація глюкози і сечовини в сироватці крові і так далі) і інші .
Якщо ми знаємо характер генетичного контролю ознаки, то ми можемо передбачити з певною ймовірністю фенотип на основі генотипу (якщо він відомий). Якщо ми не знаємо, як ознака контролюється, то ми будемо в повному невіданні, і не зможемо нічого сказати щодо ознак майбутніх поколінь. Якщо ми знаємо зв'язок між генотипом і фенотипом, то ми можемо зробити певні передбачення про розвиток ознаки (наприклад, хвороби) і, в деяких випадках, вжити заходів корисні індивіду. Для цього нам треба встановити генотип. Зараз це завдання технічно можна вирішити (оскільки секвенування генома людини проведено), хоча і непомірно дорога.
В реальності нам дано лише спостерігати прояв ознаки в поколіннях і на основі цього створювати модель генетичного контролю формування ознаки, яка може бути вірна лише в даних конкретних умовах. Але, тим не менше, якщо ми створили таку модель, то ми можемо в цьому випадку мати кошти для регуляції якоїсь ознаки, зокрема, вплинути виникнення або перебіг будь-якої хвороби. Таким чином, генетичний контроль і його вивчення має велике практичне, зокрема, медичне, значення. І в основі всього лежать Менделя закономірності, які можуть проявлятися по-різному в залежності від конкретних особливостей генотипу і середовища.
Ми зараз розглянемо, які можуть бути умови генотипу і середовища, при яких ці закономірності будуть виглядати не такими, як їх спостерігав Мендель, і чому це відбувається. Мендель спостерігав, що при об'єднанні задатків двох ознак в одному організмі може бути прояв тільки однієї ознаки. Другий задаток не проявляється. Такий тип домінування називається повним.
Відкриття законів Менделя заново призвело до виявлення інших типів домінування. Наприклад, неповного домінування, коли фенотип гетерозиготи є проміжним між двома гомозиготами. Є ще один тип домінування, дуже популярний останнім часом в суді, - кодоминирование - в гетерозиготі проявляються фенотип кожної з гетерозигот. Це явище має місце, зокрема, і у людини. Якщо у вас є татова хромосома і мамина хромосома (а це, безсумнівно, так), і вони різняться в мільйон позицій, що можна виявити різними методами, то це все випадки Кодомінування.
Первинним фенотипом організму є послідовність нуклеотидів молекул його ДНК. На цьому фенотипе будуються всі фенотипи наступних рівнів. Тобто при дослідженні вашої ДНК виявляються все і татові, і мамині ознаки; кожна з молекул ДНК проявляє свою ознаку незалежно від присутності іншої молекули ДНК з іншою ознакою: при секвенування або при розщепленні ДНК будь-якими ферментами видно обидва стану ДНК. Кодомінантність ознаки (маркери самої молекули ДНК) характеризують відмінність між хромосомами і використовуються для ідентифікації особи або встановлення батьківства (число таких випадків дозволених в суді становить кілька сотень на рік).
Коли ми говоримо про генотип і фенотип - це такі крайні єдиного процесу реалізації спадкової інформації в індивідуальному розвитку. Наприклад, гладка або зморшкувата форма горошини, це її фенотип. А генотип - це та специфічна послідовність нуклеотидів, яка в даних умовах визначає, що горошина буде гладкою або зморшкуватою. У 1999 році з менделевскими лініями гороху була проведена наступна робота. Ділянки хромосоми, що відповідають за форму горошини, були клоновані, секвенувати, і було встановлено їх особливості - відмінність послідовностей нуклеотидів - які і визначали розвиток гладкою або зморшкуватою форми горошини.
Зверніть увагу, що форма горошини є кінцевим ознакою, а формування ознаки цього рівня передує прояв ознаки на багатьох попередніх рівнях. По-перше, це наявність (аллель 1) або відсутність (аллель 2) олігосахариду, яке і призводить до тієї або іншій формі горошини. Ще більш глибоким рівнем прояви фенотипу є те, що є відповідний білок (аллель 1), який необхідний для синтезу олігосахариду або це той же білок, але альтернативної структури (аллель 2), при якій олігосахарид не утворюється. Ще більш глибоким ознакою є РНК, з якою синтезується цей білок. Ці РНК різні за послідовності нуклеотидів (аллели 1 і 2), що і робить різними відповідні білки. А ці РНК різні, тому що транскрибується з різних молекул ДНК, татової і маминої, у яких послідовності нуклеотидів в даній позиції різні (аллели 1 і 2). Все це - прояв одного і того ж фенотипу, послідовно реалізується на кожному з рівнів.
Ми маємо право говорити про фенотипе на кожному з цих багатьох рівнів - від конкретних особливостей послідовності нуклеотидів ДНК до форми горошини. При цьому, як тільки ми просуваємося від ДНК вище, тим більше вплив умов навколишнього середовища. Наприклад, можливість функціонування різних алелей на рівні ДНК (транскрипція татової і маминої копії гена) буде мало залежати від температури, а можливість функціонування тих же алелей на рівні білка може критично залежати від температури. При деяких температурах білок (наприклад, аллель 1) буде працювати, а інший (аллель 2) не буде працювати. Як тільки ми висуваємося на більш високий рівень в реалізації фенотипу, з'являється більше можливостей для впливу навколишнього середовища на прояв ознаки.
І навпаки, чим ближче ми просуваємося до генотипу, тим передбачувані зв'язок між генотипом і фенотипом.