Генетика - наука про спадковість і мінливість. Основні генетичні терміни і поняття

Наука про спадковість і мінливість організмів. Перші спроби дослідження спадковості відносяться до 17 ст. У 1694 р ньому. ботанік Р. Камераріус довів існування статевих відмінностей у рослин. На початку 18 ст. були отримані штучні гібриди рослин. У 1760 р член Петербурзької академії наук І. Кельрейтер встановив успадкування гібридами ознак батьків і довів «рівноправність» чоловічих і жіночих клітин у формуванні зав'язі, а також запропонував методи штучного схрещування, к-які використовуються до теперішнього часу. З початку 19 ст. робилися численні спроби з'ясувати закони успадкування нащадками ознак батьків. Зробити це вдалося лише в другій поло-. вини 19 в. чещ. натуралісту І. Г. Менделя. У 1865 р він доповів результати своїх багаторічних спостережень. Вперше застосувавши статистичні методи обробки результатів Біологічний. експериментів, І. Г. Мендель сформулював основні закони передачі спадкових ознак від батьків до нащадків (правила або закони Менделя - см. Спадковість). Більш того, І. Г. Мендель висловив припущення про існування в клітинах окремих (дискретних) частинок, які є задаткам !! спадкових ознак. Кожна соматична клітина (клітина тіла) несе пару спадкових задатків, в статевих клітинах (гаметах) міститься по одному завдатку з пари. При заплідненні, коли відбувається злиття статевих клітин, ці задатки об'єднуються в різних комбінаціях. Їх прояв у процесі формування нового організму лежить в основі відтворення спадкових ознак батьків. В умовах, коли науці ще не були відомі багато суттєві деталі будови клітини, клітинний розподіл, сутність статевого процесу, коли уявлення про спадковість носили умоглядний характер, геніальний здогад І. Г. Менделя, заснована лише на бездоганно точних дослідах, їх математичної обробки і глибокому логічному аналізі причин спостережуваних явищ, настільки випереджала свою епоху, що не могла бути зрозуміла і оцінена сучасниками. Тільки через 35 років одночасно і незалежно один від одного голл. вчений Г. де Фриз, ньому. вчений К. Корренс і чеськ. вчений Е. Чермак вдруге «відкрили» закони успадкування ознак і, випадково виявивши забуту роботу І. Г. Менделя, познайомили з нею наукову громадськість. З цього часу вчення про спадковість почав розвиватися як самостійна наука, к-раю з 1906 р за пропозицією англ. біолога У. Бетсона стала іменуватися генетикою. Спадкові задатки дат. вчений У. Иоганнсен в 1909 році запропонував називати генами.
Початок 20 ст. ознаменувався бурхливим розвитком генетичних досліджень. До цього часу завдяки успіхам мікроскопіч. техніки стали відомі деталі будови клітини, були відкриті закономірності утворення статевих клітин, виявлені хромосоми - особливі структури в ядрі клітини, число і набір яких брало виявилися досить стабільні в клітинах організмів одного виду і різні в клітинах організмів різних видів; було відкрито розподіл клітин - мітоз, під час к-якого відбувається точний розподіл подвоїти хромосом батьківських клітин по дочірнім клітинам. Тому саме хромосоми стали вважати носіями спадкових задатків - генів. З'ясування закономірностей дозрівання статевих клітин і сутності статевого процесу в цілому повністю підтвердили припущення І. Г. Менделя про дискретності і парності спадкових задатків. "З цих пір явищеспадковості стали міцно зв'язувати з матеріальними структурами клітини, якими є хромосоми.
У 1899-1901 рр. голл. вчений Г. де Фриз і рус. вчений С. І. Коржинський виявили організми, що різко відрізняються від своїх родичів по будь-якою ознакою, і сформулювали теорію, згідно якої спадкові властивості і ознаки організму можуть раптово і різко змінюватися. Ця теорія отримала назву «теорії мутацій» (див. Мінливість, Мутагенез), а особини, що різко відрізняються по будь-якою ознакою від своїх родичів, були названі мутантами.
У 1910-1913 рр. амер. біолог Т. Морган і його учні в дослідах на плодової мушки дрозофіли довели, що гени дійсно зосереджені в хромосомах в лінійному порядку; був експериментально встановлений процес обміну ділянками хромосом під час зближення парних хромосом і перехрещення між ними (кросинговеру). Слідом за цим були побудовані перші карти хромосом спочатку для широко використовуваного в генетичних експериментах комахи дрозофіли, а потім і для ряду інших організмів (перш за все корисних рослин). На картах вказувалося взаємне розташування генів і відносне відстань між ними. Ця робота стала можливою, зокрема, тому, що Т. Моргану і його учням вдалося виявити в природних умовах мутантних дрозофіл по кільком сотням генів. Вчення Т. Моргана, що отримало назву хромосомної теорії спадковості, стало одним з найбільш великих материалистич. узагальнень молодий Г. на початку 20 ст.
У той же час в методологія, плані вивчення розташування генів в хромосомах, що проводилося школою Т. Моргана, і аналіз комбінування генів при схрещуванні різних організмів велися окремо. Вчені не бачили на перших порах чітких зв'язків між обома напрямками генетичних досліджень. Чи не сприяло їх об'єднанню і вивчення мутацій і частоти виникнення їх в природних умовах. Більш того, обидва засновника мутаційної теорії (і Г. де Фриз, і СІ. Коржинський) протиставляли процес виникнення мутацій вченню Ч. Дарвіна (див. Еволюційне учення). Кілька по-різному в генетичному сенсі, але в методоло-гич. плані фактично однаково С. І. Коржинський і Г. де Фриз навіть намагалися замінити дарвінізм мутаційної теорією, вважаючи, що основним фактором еволюції живої природи є не процес природного відбору, т. е. виживання найбільш пристосованих до умов існування форм, а процес мимовільного виникнення мутантних форм організмів. Їх погляди були підхоплені низкою апологетів їх теорії, і прірва, разверзшаяся між вченням І. Г. Менделя (пізніше і Т. Моргана) н дарвінізмом, поглиблювалася все більше. Тим більше значним внеском у розвиток генетики і еволюційного вчення стала теоретична робота сов. вченого С. С. Четверикова, який довів в 1926 р, що саме мутації, що виникають в природних умовах і підкоряються спадкоємства за закономірностям І. Г. Менделя, служать основним матеріалом для природного добору. С. С. Четвериков математично розрахував швидкість відбору і розподілу мутантних генів в ряду поколінь організмів в популяціях і тим самим заклав основи найважливішої галузі Г. по-пуляціонной генетики. Велику роль для розуміння можливостей мутаційного процесу і типів виникаючих мутацій зіграв сформульований сов. вченим М. І. Вавілов закон гомологічних рядів в спадкової мінливості. Сутність цього закону полягає в тому, що виникають нові спадкові ознаки (мутації) у родинних організмів (видів, пологів і сімейств) однотипні (паралельні) і ці ознаки повторюють один одного тим повніше, чим ближче зіставляються форми за своїм походженням. Н. І. Вавилов провів величезну роботу по обстеженню центрів походження культурних рослин, пошуку по всьому світу стародавніх предків сучасних культурних рослин, різних змінених форм цих рослин (мутантів), створення всеосяжної колекції світових рослинних ресурсів і, нарешті, створення вчення про теоретичні основи селекції рослин.
У генетичних дослідженнях, проведених до 1925 р, вчені використовували мутанти, що зустрічаються в природних умовах. Хоча ряд вчених (Т. Морган, Н. К. Кольцов і ін.) Розуміли, що можна викликати мутації штучно, численні спроби здійснити це експериментально довгий час виявлялися безуспішними. Лише в кінці 20-х - початку 30-х рр. була експериментально доведена можливість мінливості генів (мутації) під впливом чинників навколишнього середовища. У 1925 р сов. вчені Г. А. Надсон і його учень Г. С. Філіппов на нижчих дріжджах, а в 1927 р амер. вчений Г. Меллер на дрозофілі отримали спадкові зміни (мутації) під дією рентгенівських променів; в 1928 р інший учень Г. А. Над-сона - М. Н. Мейсель показав здатність хімічних. агентів викликати мутації у дріжджів. У 1932 р явище химич. мутагенезу у дрозофіли спостерігав сов. вчений В. В. Сахаров, а починаючи з 1939 р сов. генетик І. А. Рапопорт почав широке вивчення мутагенної (викликає мутації) активності багатьох хімічних сполук. Великий внесок у вивчення цієї проблеми внесли англ. вчені Ш. Ауербах, Т. Лавли і ін.
В результаті всіх цих досліджень вже в кінці 20-х рр. перед генетиками особливо гостро постало питання, що ж являє собою ген як структурна одиниця спадковості і яка, його хімічні. природа.
Спроби знайти відповідь на перше питання були зроблені ще Т. Морганом, а також рядом інших дослідників (в т. Ч. Сов. Вченими А. С. Срібний,-ровськ, Н. В. Тимофєєвим-Ресовський, А. А. Прокоф-Бельговской в співавторстві з Р. Меллером, який приїхав на кілька років працювати в СРСР, і ін.). Було встановлено, що кожен ген визначає розвиток певної ознаки і є мінімальною частиною хромосоми, к-раю може бути передана в іншу хромосому в процесі їх перехрещення (кросинговеру). Вважалося також встановленим, що в результаті ген змінюється цілими, не дроблячись. Однак в 1928-1929 рр. в лабораторії А. С. Серебровского було доведено, що в певних умовах вдається розділити ген на різні ділянки (центри), мутують окремо, і в подальшому їм і його учнями, перш за все Н. П. Дубиніним, була сформульована так зв. центрова теорія гена. Сутність цієї теорії полягала в тому, що ген складається з окремих розташованих в лінійному порядку частин, що окремі частини гена можуть незалежно один від одного змінюватися (мутувати), обмінюватися при перехрестя хромосом (кроссинговере) і що дія гена в цілому обумовлено об'єднанням і узгодженням функцій його частин. Пізніше, в 1957-1961 рр., Амер. вчений С. Бензер експериментально довів, що по довжині гена може виникати безліч мутацій і що при кроссннговере в обміні беруть участь не цілі гени, а їх окремі ділянки.
Складніше було з вивченням хім. природи генів. Ще в другій половині 19 ст. було з'ясовано, що в хромосомах містяться білки і нуклеїнові кислоти і що одна з нуклеїнових к-т - дезоксірібонуклеі-нова к-та (ДНК) міститься гл. обр. в хромосомах. Однак біологи аж до 50-х рр. 20 в. вважали, що гени повинні міститися в молекулах білків. Лише після зародження нового напрямку в Г. молекулярної генетики вдалося довести, що саме ДНК несе генетичну функцію. Перші експерименти, прямо показали це, були зроблені на мікроорганізмах. У 1944 р було встановлено, що введення ДНК, взятої від одних бактерій, в клітини інших викликає у останніх зміни спадкових властивостей. У 1953 р амер. вчений Дж. Уотсон і англ. вчений Ф. Крик запропонували гіпотезу про будову ДНК, згідно якої молекула ДНК має форму подвійної спіралі. Гіпотеза Дж. Уотсона і Ф. Кріка, швидко отримала підтвердження, визнання і розвиток, лежить в основі сучасних уявлень про будову і властивості нуклеїнових к-т. Відносна хім. стабільність і присутність ДНК і РНК у складі всіх без винятку живих організмів дозволили припустити, що саме ДНК і РНК, а не білки є тими молекулярними структурами, к-які забезпечують зберігання і передачу генетичної інформації - успадкування ознак від клітини до клітини і через статеві клітини від організму батьків до нащадків. Іншими словами, саме ДНК і РНК містять у своїй структурі «запис» спадкової інформації, той генетичний код, ту спадкову програму, згідно якої при розвитку нового організму будуть формуватися його ознаки та властивості. Гіпотеза Уотсона - Кріка давала можливість зрозуміти, як відбувається постійне і точне відтворення молекул ДНК (реплікація), т. Е. Як забезпечується збереження і підтримання генетичної програми. Сама ідея про можливість відтворення спадкового матеріалу була висловлена сов. вченим Н. К. Кольцовим ще в 1927 р Однак Н. К. Кольцов пов'язував цю здатність з функцією білкових молекул хромосом. Надалі було встановлено, що відтворення (реплікація) молекул ДНК відбувається шляхом подвоєння її ланцюгів попередніми роз'єднанням подвійної спіралі на дві одиночні і шляхом добудови на кожній з них її копії відповідно до правила соответственности, комплементарності (див. Генетичний код). Подальші дослідження розкрили складний механізм відтворення ДНК, виявили роль ферментів в цьому процесі, показали, що принципова схема реплікації подібна як у нижчих, так і у вищих організмів (див. Молекулярна генетика, Нуклеїнові кислоти).
Паралельно з цим почала вимальовуватися картина функціонування спадкової записи, т. Е. Здійснення генетичного контролю синтезу білків. З'ясувалася роль в цьому процесі молекул двох видів РНК - так зв. інформаційної РНК і транспортної РНК. Можливості молекулярного аналізу структур ядра клітини озброїли Г. новими точними, експериментально доведеними даними про структуру і властивості генів. Було розвинене уявлення про гені як відрізку молекули ДНК (у деяких вірусів РНК), к-рий визначає будову первинної структури молекули білка, є основою сучасної Г. (див. Ген, Генетичний код).
Подальшим кроком в пізнанні функції генів було встановлення механізмів регуляції їх роботи. У 1961 р франц. вчені Ф. Жакоб і Ж. Моно виявили, що у мікроорганізмів поряд з генами, що визначають синтез ферментів (так зв. структурними генами), існують ділянки ДНК, що керують активністю цих структурних генів, - так зв. регуляторні ділянки. Було з'ясовано, що є гени-регулятори, які кодують білки-репрессори, останні «закривають» або, навпаки, «відкривають» операторні ділянки, після чого стає можливим з'єднання ферментів, що ведуть синтез копій генів у вигляді інформаційних РНК (РНК-полімерази) з молекулами ДНК. Просуваючись по гену, РНК-полімераза веде копіювання (зчитування, або транскрипцію) генетичної інформації.
Починаючи з 1960 р стали проводитися дослідження молекулярних основ виникнення мутацій, в результаті яких брало була виявлена загальна схема змін генетичного коду, що призводять до мутацій. Після откритія.у мікроорганізмів тварин і рослин властивості відновлення (репарації) генетичних ушкоджень у результаті роботи спеціальних ферментів (див. Репарація генетична) стали накопичуватися дані про зв'язок мутагенезу і репарації. В ході цих досліджень було з'ясовано, що пошкодження генів системи репарації призводить до ряду серйозних захворювань. Було встановлено, що частота хромосомних порушень прямо залежить від рівня активності репаруючу систем. Т. о., Ці відкриття, зроблені в найкоротші терміни, дозволили створити струнку матеріалістичну концепцію пристрою і функціонування генетичного апарату живих клітин.
Однак слід підкреслити, що
переважна більшість досліджень
ний, присвячених вивченню генетичних
ського коду, було проведено на мікро
організмах і ще належить з'ясувати,
наскільки справедлива концепція регу
ляции дії генів, прийнята для
бактерій і вірусів, щодо більш
високоорганізованих організмів.
Сов. вчені А. С. Спірін та Г. П. Георгієв (1964-1965) експериментально довели, що в клітинах тварин існують особливі форми збереження генетичної інформації у вигляді сформованих раніше на молекулах ДНК комплексів РНК і білка (так зв. інформосоми). У лабораторії Г. П. Георгієва були проведені широкі дослідження структури ДНК і характеру зчитування (транскрипції) генетичної інформації в хромосомах вищих організмів. У 1959 р амер. вчені Бріттен і Кон виявили, що у вищих організмів, на відміну від мікробів, в ДНК хромосом є області, в яких брало послідовність розташування нуклеотидів (основних структурних одиниць нуклеїнових к-т, і зокрема ДНК) багаторазово повторена. Роль цих повторів поки ще не ясна, але встановлено, що ці зони відмежовують області так зв. унікальних послідовностей, в яких брало, мабуть, і зосереджені структурні гени. У 1974-1976 рр. став прояснюватися характер взаємодії ДНК і особливих білкових молекул (гістонів) і склалося уявлення про організацію хромосом на молекулярному рівні (див. Хромосоми).
Великих успіхів досягнуто у вивченні Г. соматичних клітин (клітин тіла) людини, тварин і рослин.
Одним з найбільш захоплюючих і найменш з'ясованих питань Г. є питання про генетичне контролі розвитку, про реалізацію програми розвитку, закодованої в генах, в сукупність ознак і властивостей організму (так зв. Біологія розвитку). Питання це, природно, включає в себе і проблему співвідношення спадкового та середовищного факторів у формуванні організму.
З'ясування молекулярних механізмів реалізації генетичної програми, т. Е. Дані молекулярної Г., сприяли переходу на новий рівень досліджень в області вивчення індивідуального розвитку - ембріології, вікової морфології і фізіології, геронтології (наука про старіння організму), а в медицині - в області акушерства , педіатрії та інших розділів клинич. медицини. Центральним стало питання про інтимні механізми диференціювання клітин, зародка, в результаті яких брало, напр., За кілька місяців з заплідненої жіночої статевої клітини (зиготи) розвивається складний багатоклітинний організм людини з різноманітними і високорозвиненими функціональними можливостями.
Старий питання про роль спадковості і факторів навколишнього середовища у формуванні та розвитку організму після багатьох років значних протиріч, дискусій і помилок дозволяється на базі визнання єдності генетичного і середовищного факторів. В процесі розвитку кожного організму реалізація генетичної програми в якості і ознаки нового організму йде під впливом оточуючих організм умов існування. Наявність генетичної програми забезпечує успадкування організмом ознак предків, його видових особливостей, а вплив умов існування в період розвитку організму визначає відхилення, індивідуальні відмінності, к-які, як правило, не зачіпають генетичного апарату статевих клітин і не успадковуються. Якщо ж під впливом сильнодіючих факторів середовища виникла мутація, то така зміна успадковується і може зберегтися в поколіннях. Сучасна Г. залежно від конкретних об'єктів, завдань і методів дослідження має ряд розділів і напрямків. В рамках Г. сформувалися як самостійні наукові дисципліни молекулярна генетика, біохімічн. генетика, генетика розвитку, цитогенетика, радіаційна генетика, популяційна-ва генетика, генетика мікроорганізмів (див. Бактерії, Віруси) і ін. Спеціалізація всередині кожної з цих дисциплін досягла настільки великій мірі, що багато хто з них сприймаються на перший погляд як самостійні, що не пов'язані один з одним галузі науки. Разом з тим, незважаючи на гадану ізольованість, кожна з цих дисциплін має на меті пізнання різних сторін таких фундаментальних властивостей живого, як спадковість і мінливість.
Така загальна методологич. спрямованість досліджень різних сторін прояву общебіологіч. властивостей спадковості і мінливості, безсумнівно, призведе в майбутньому до об'єднання всіх поки різнорідних і часом навіть зовні не пов'язаних генетичних дисциплін в єдину науку. Цей процес проявляється, звичайно, і зараз, але він ще далекий від завершення.
Дедалі більшого розвитку отримують генетичні дослідження, спрямовані на пізнання спадковості людини. Велика увага приділяється вивченню генетичних основ спадкових хвороб, з'ясування ролі чинників довкілля в розвитку хвороб і значення охорони середовища як потужного чинника, що впливає на спадковість людини. Ці питання вирішує спеціальний розділ Г. медична генетика.
В епоху науково-технічної революції Г. є одним з найбільш актуальних бурхливо розвиваються розділів біології, завжди тісно пов'язаним з практикою. Саме успіхи сучасної Г. сприяють збільшенню продуктивності с.-г. культур (пшениці, цукрових буряків, кукурудзи, гречки та ін.), отримання кращих порід худоби, хутрових звірів. Досягнення Г. сприяють розвитку мікробіологіч. промисловості (виробництво антибіотиків і ін.) - Успіхи сучасної Г. дозволяють підійти до грандіозної задачі майбутнього - розробці заходів захисту спадкового апарату людини від шкідливих впливів факторів навколишнього середовища, а також способів лікарської корекції (виправлення) спадкових недуг і можливого в майбутньому прямого втручання в спадковий апарат людини з метою усунення або заміни «хворих» генів, від яких брало залежить те чи інше захворювання. В цьому плані висловлюються надії, "то використання спеціальних MeTOxici біохімічн. Маніпулювання з генами (див. Інженерія генетична) дозволить в майбутньому звільнити людину від вантажу багатьох хвороб. Однак очевидно, що до цього часу треба буде розв'язати величезну кількість як суто наукових, так і не менш важливих морально-етичних проблем. про основні поняття і закони вчення про спадковість і мінливість в сучасному їх розумінні - см. ген , Мінливість, Мутагенез, Спадковість
Основні терміни і поняття, що перевіряються в екзаменаційній роботі: алельних гени, аналізує схрещування, взаємодію генів, ген, генотип, гетерозиготность, гіпотеза чистоти гамет, гомозиготность, дигибридное схрещування, закони Г. Менделя, кількісні ознаки, Кроссинговер, літали, множинні алелі, моногибридное схрещування, Незалежне успадкування, неповне домінування, правило одноманітності, розщеплення, фенотип, цитологічні основи законів Менделя.
генетика - наука про спадковість і мінливість організмів. Ці дві властивості нерозривно пов'язані один з одним, хоча мають протилежну спрямованість. Спадковість передбачає збереження інформації, а мінливість цю інформацію змінює. спадковість - це властивість організму повторювати в ряді поколінь свої ознаки і особливості свого розвитку. Мінливість - властивість організмів змінювати свої ознаки під впливом зовнішнього або внутрішнього середовища, а також в результаті нових генетичних комбінацій, що виникають при статевому розмноженні. Роль мінливості полягає в тому, що вона «поставляє» нові генетичні комбінації, що піддаються дії природного відбору, а спадковість зберігає ці комбінації.
До основних генетичним поняттям ставляться такі:
ген - ділянку молекули ДНК, в якому закодована інформація про послідовність амінокислот у молекулі білка.
аллель - пара генів, що відповідають за альтернативне (різне) прояв одного і того ж ознаки. Наприклад, за колір очей відповідають два алельних гена, розташованих в однакових локусах (місцях) гомологічних хромосом. Тільки один з них може відповідати за розвиток карих лаз, а інший - за розвиток блакитних очей. У тому випадку, коли обидва гени відповідають за однаковий розвиток ознаки, говорять про гомозиготном організмі за цією ознакою. Якщо алельних гени визначають різне розвиток ознаки, говорять про гетерозиготному організмі.
Алельних гени можуть бути домінантними , Що пригнічують альтернативний ген, і рецесивними , Придушуваними.
Сукупність генів організму називається генотипом даного організму. Генотип організму описується словами - «гомозиготний» або «гетерозиготний». Однак не всі гени проявляються. Сукупність зовнішніх ознак організму називається його фенотипом. Кароокий, повний, високий - це спосіб опису фенотипу організму. Кажуть також про домінантному або рецесивним фенотипе.
Генетика вивчає закономірності успадкування ознак. Основним методом генетики єгібрідологіческій метод або схрещування. Цей метод був розроблений австрійським вченим Грегором Менделем в 1865 р
Розвиток генетики спричинило за собою розвиток багатьох наукових напрямків і, перш за все, еволюційного вчення, селекції рослин і тварин, медицини, біотехнології, фармакології та ін.
На рубежі XX і XXI століть розшифрований геном людини. Вчених вразило, що у нас всього 35 000 генів, а не 100 000, як думали раніше. У круглого хробака 19 тис. Генів, у гірчиці - 25 тис. Відмінності між людиною і шимпанзе становлять 1% генів, а з мишею - 10%. Людині дісталися в спадок і гени, яким 3 мільярди років і відносно молоді гени.
Що дає науці прочитання генома? Перш за все, це знання дозволяє цілеспрямовано вести генетичні дослідження по виявленню як патологічних, так і потрібних, корисних генів. Вчені не полишають надії на одужання людей від таких захворювань, як рак і СНІД, діабет і ін. Також не залишають надії і на подолання старезної старості, передчасної смертності та багатьох інших бід людства.
Закономірності спадковості, їх цитологічні основи. Моно- і дигибридное схрещування. Закономірності успадкування, встановлені Г. Менделем. Зчеплене успадкування ознак, порушення зчеплення генів. Закони Т. Моргана. Хромосомна теорія спадковості. Генетика статі. Успадкування ознак, зчеплених зі статтю. Генотип як цілісна система. Розвиток знань про генотипі. Геном людини. Взаємодія генів. Рішення генетичних задач. Складання схем схрещування. Закони Г. Менделя і їх цитологічні основи
Терміни і поняття, що перевіряються в екзаменаційній роботі: алельних гени, аналізує схрещування, ген, генотип, гетерозиготность, гіпотеза чистоти гамет, гомозиготность, дигибридное схрещування, закони Менделя, моногибридное схрещування, морганіда, спадковість, незалежне успадкування, неповне домінування, правило одноманітності, розщеплення, фенотип, хромосомна теорія спадковості, цитологічні основи законів Менделя.
Успіх роботи Грегора Менделя був пов'язаний з тим, що він правильно вибрав об'єкт дослідження і дотримувався принципи, які є основою гибридологического методу:
1. Об'єктом дослідження стали рослини гороху, що належали до одного виду.
2. Досвідчені рослини чітко відрізнялися за своїми ознаками - високі - низькі, з жовтими і зеленими насінням, з гладкими і зморшкуватими насінням.
3. Перше покоління від вихідних батьківських форм завжди було однаковим. Високі батьки давали високу потомство, низькі батьки давали рослини маленького росту. Таким чином, вихідні сорти були так звані «чисті лінії».
4. Г. Мендель вів кількісний облік нащадків другого і наступних поколінь, у яких спостерігалося розщеплення в ознаках.
Закони Г. Менделя описують характер успадкування окремих ознак протягом декількох поколінь.
Перший закон Менделя або правило одноманітності.Закон виведений на основі статистичних даних, отриманих Г. Менделем при схрещуванні різних сортів гороху, які мали чіткі альтернативні відмінності за такими ознаками:
- форма насіння (кругла / некругла);
- забарвлення насіння (жовта / зелена);
- шкірка насіння (гладка / зморшкувата) і т.д.
При схрещуванні рослин з жовтими і зеленими насінням Мендель виявив, що всі гібриди першого покоління виявилися з жовтими насінням. Він назвав цей ознака домінантним. Ознака, що визначає зелене забарвлення насіння, був названий рецесивним (відступаючим, пригніченим).
Так екзаменаційна робота вимагає від учнів уміння правильно оформляти записи при вирішенні генетичних завдань, то ми покажемо приклад такого запису.
1. На підставі отриманих результатів і їх аналізу Мендель сформулював свій перший закон . При схрещуванні гомозиготних особин, які відрізняються однією або декількома парами альтернативних ознак, Все гібриди першого покоління виявляться за цими ознаками одноманітно і схожими на батька з домінантною ознакою.
В разі неповного домінування тільки 25% особин фенотипически схожі на батька з домінантною ознакою і 25% особин будуть схожі на рецесивного по фен-типу батька. Решта 50% гетерозигот будуть від них фенотипически відрізнятися. Наприклад, від красноцветковими і белоцветкових рослин лев'ячого зіва в потомстві 25% особин червоні, 25% - білі, а 50% - рожеві.
2. Для виявлення гетерозиготности особини за певним алелем, тобто наявності рецесивного гена в генотипі, використовується аналізує схрещування . Для цього особина з домінантною ознакою (АА? Або Аа?) Схрещують з гомозиготною по рецесивним аллели особиною. У разі гетерозиготності особини з домінантною ознакою розщеплення в потомстві буде 1: 1
АА × аа → 100% Аа
Аа × аа → 50% Аа і 50% аа
Другий закон Менделя або закон розщеплення.При схрещуванні гетерозиготних гібридів першого покоління між собою, у другому поколінні виявляється розщеплення за цією ознакою. Це розщеплення носить закономірний статистичний характер: 3: 1 за фенотипом і 1: 2: 1 за генотипом. У разі схрещування форм з жовтими і зеленими насінням відповідно до другого законом Менделя отримують такі результати схрещування.
З'являються насіння як з жовтою, так і з зеленим забарвленням.
Третій закон Менделя або закон незалежного успадкування при дигибридном (полигибридном) схрещуванні.Цей закон виведений на основі аналізу результатів, отриманих при схрещуванні особин, що відрізняються по двох парах альтернативних ознак. Наприклад: рослина, що дає жовті, гладкі насіння схрещується з рослиною, що дає зелені, зморшкуваті насіння .
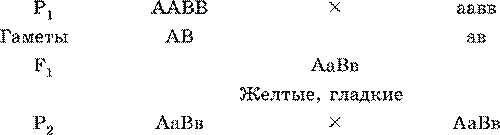
Для подальшої записи використовуються ґрати Пеннета:

У другому поколінні можлива поява 4 фенотипів щодо 9: 3: 3: 1 і 9 генотипів.
В результаті проведеного аналізу з'ясувалося, що гени різних алельних пар і відповідні їм ознаки передаються незалежно один від одного. Цей закон справедливий:
- для диплоїдних організмів;
- для генів, розташованих в різних гомологічних хромосомах;
- при незалежному розбіжності гомологічних хромосом в мейозі і їх випадковому поєднанні при заплідненні.
Зазначені умови і є цитологическими основами дигибридного схрещування.
Ті ж закономірності розповсюджуються на полигибридного схрещування.
В експериментах Менделя встановлена дискретність (переривчастість) спадкового матеріалу, що пізніше призвело до відкриття генів, як елементарних матеріальних носіїв спадкової інформації.
Відповідно до гіпотези чистоти гамет в сперматозоїді або яйцеклітини в нормі завжди знаходиться тільки одна з гомологічних хромосом даної пари. Саме тому при заплідненні відновлюється диплоїдний набір хромосом даного організму. розщеплення - це результат випадкового поєднання гамет, що несуть різні алелі.
Так як події випадкові, то закономірність носить статистичний характер, тобто визначається великим числом рівноймовірно подій - зустрічей гамет, що несуть різні (або однакові) альтернативні гени.
Приклади ЗАВДАНЬ
1. Етапи розвитку ГЕНЕТИКИ ЯК НАУКИ.
генетика- наука про спадковість і мінливість організмів, про закономірності спадкової мінливості і про матеріальні основи спадковості.
а) Розвиток класичної генетики (створення самої науки)
IV-V століття до н. е. - перші згадки в релігійній літературі в зв'язку з сільськогосподарської та медичної діяльністю.
1859 г. - «Походження видів шляхом природного відбору» (Чарльз Дарвін)
1865 р - «Досліди над рослинними гібридами» (Грегор Мендель, після виступу в м Брно)
На початку XX століття роботи Менделя знову привернули увагу в зв'язку з дослідженнями Гуго Де Фріза (Голландія), Карла Корренса (Німеччина) і Еріха фон Чермака (Австрія) по гібридизації рослин, в яких були підтверджені основні висновки про незалежне успадкування ознак і про численних співвідношеннях при «розщепленні» ознак в потомстві.
=\u003e Відкриття і перевідкриття законів Менделя. «Тріумфальна хода» менделізму.
Незабаром англійський натураліст Вільям Бетсон ввів у вживання назву нової наукової дисципліни: генетика (в 1905 р в приватному листі і в 1906 р публічно).
б) Вивчення спадкових явищ
1901-1903 рр. де Фриз сформулював мутационную теорію.
Основні положення мутаційної теорії:
1. Мутації раптові, як дискретні зміни ознак
2. Нові форми стійкі
3. На відміну від спадкових змін, мутації не утворюють безперервних рядів, що не групуються навколо будь-якого середнього типу. Вони являють собою якісні скачки змін
4. Мутації проявляються по-різному і можуть бути як корисними, так і шкідливими
5. Ймовірність виявлення мутацій залежить від числа досліджуваних особин
6. Подібні мутації можуть виникати неодноразово
Хромосомна теорія спадковості була сформульована в 1900-1903 рр. американським цитологом Вільямом Сеттона і німецьким ембріологів Теодором Бовери і надалі розвинена знаменитим американським генетиком
Томасом Морганом
Хромосомна теорія спадковості - теорія, згідно з якою хромосоми, укладені в ядрі клітини, є носіями генів і являють собою матеріальну основу спадковості
в) Вивчення на молекулярному рівні
1928 р.- "Хромосома являє собою макромолекулу (білок), окремі радикали якого виконують різні функції генів" - Н. К. Кольцов (видатний російський біолог)
1929-1931 рр. А. Серебровский - складна структура гена (В кінці 1920-х років радянські генетики А. С. Серебровський і Н. П. Дубінін експериментально показали, що ген не є одиницею мутації, що він має складну структуру: складається з декількох субодиниць, здатних самостійно мутувати)
40-ті роки - Джордж Бідл і Едвард Татум узагальнили зв'язок між ферментом і геном у вигляді постулату один фермент - один ген. (Оскільки ферменти-це білки, а багато білків складаються з більш ніж одного типу поліпептидних ланцюгів, постулат в подальшому став формулюватися як «один поліпептид - один ген».)
1944 р О. Евері, К.Мак-Леод і М.Мак-Карті (США) прийшли до висновку, що ДНК може бути носієм генетичної інформації.
1953 г. - Дж.Уотсон і Ф. Крик (США) побудували просторову модель молекули дезоксирибонуклеїнової кислоти (ДНК). Вони визначили, що молекула ДНК складається з двох ланцюгів, закручених одна навколо іншої (подвійна спіраль).
У 1968 Маршалл Уоррен Ніренберг, Роберт Вільям Холлі і Хар Гобінд Корану отримали Нобелівську премію «за розшифровку генетичного коду і його функції в синтезі білків».
Р. Фішер, Дж. Холдейн і Сьюелл Райт (20-30-і рр.) Отримали основні популяційно-генетичні слідства із законів Менделя. (Були закладені основи генетико-математичних методів вивчення процесів, що відбуваються в популяціях.)
1920 р Фундаментальний внесок у генетику популяцій вніс С. С. Четвериков (1926), який об'єднав в єдиній концепції закономірності менделізму і дарвінізму. (=\u003e Синтетична теорія еволюції)
2. ОСНОВНІ НАПРЯМКИ СУЧАСНОЇ ГЕНЕТИКИ.
Основні напрямки сучасної генетики - вивчення передачі спадковості на молекулярному рівні і вплив на передачу спадковості. Сучасна генетика ділиться на генетику рослин, тварин і людини.
1. Генетика домашніх рослин і тварин також іменується селекцією. Селекція вивчає методи поліпшення продуктивності в сільському господарстві.
2. Стабільно розвивається генетичне відтворення тварин і людини, минаючи статеве розмноження - клонування. Клонування це народження повноцінної особини тварини або людини з не статевий клітини особини батька. На відміну від статевого розмноження клонування не припускав мінливості.
3. Генетика людини отримала своє значення для науки і медицини порівняно недавно. Розшифровка геному людини дала можливість вивчати і впливати на передачу недоброякісних генів від батьків до дітей.
3. ОСНОВНІ ПОНЯТТЯ І ТЕРМІНИ СУЧАСНОЇ ГЕНЕТИКИ.
Спадковість - властивість організмів забезпечувати матеріальну і функціональну наступність між поколіннями, а також повторювати певний тип індивідуального розвитку.
Мінливість - варіабельність (різноманітність) ознак серед представників даного виду.
Спадкування - передача генетичної інформації одним поколінням іншому (батьками або батьком - потомству).
Наследуемость - частка фенотипической мінливості в популяції, обумовлена генетичною мінливістю (відповідно до зазначеного якісному або кількісному ознакою).
Ген - послідовність нуклеотидів в ДНК, яка кодує певну РНК.
Геном - загальна генетична інформація, що міститься в генах організму, або генетичний склад клітини.
Генотип - 1) вся генетична інформація організму; 2) генетична характеристика організму по одному або декільком досліджуваним локусам.
Фенотіп- зовнішній прояв властивостей організму, що залежать від його генотипу і факторів навколишнього середовища.
Алельних гени - різні форми одного і того ж гена, розташовані в однакових ділянках (локусах) гомологічних хромосом. Аллели визначають варіанти розвитку одного і того ж ознаки.
Гомозиготний організм - організм, який має дві ідентичні гамети даного гена в гомологічних хромосомах.
Гетерозиготний - різні гамети.
Норма реакції - 1) межа модифікаційної мінливості ознаки, обумовлений генотипом. Зазвичай якісні ознаки мають вузькі межі мінливості, а кількісні ознаки змінюються в широких пределах.2) здатність генотипу формувати в онтогенезі, залежно від умов середовища, різні фенотипи.
Альтернативні ознаки - взаємовиключні, контрастні ознаки.
Домінантний аллель - виражена в фенотипі незалежно від присутності в геномі іншого аллеля.
Рецесивні аллели - придушуються домінантними.
4. ЕВОЛЮЦІЯ ПОНЯТТЯ «ГЕН».
СТРУКТУРНО-ФУНКЦІОНАЛЬНА ОРГАНІЗАЦІЯ ГЕНА, ЙОГО ХІМІЧНА ПРИРОДА.
Ген - структурна і функціональна одиниця спадковості, що контролює розвиток певної ознаки чи властивості. Сукупність генів батьки передають нащадкам під час розмноження.
Спочатку термін ген з'явився як теоретична одиниця передачі дискретної спадкової інформації.
В даний час, в молекулярної біології встановлено, що гени - це ділянки ДНК, що несуть якусь цілісну інформацію - про будову однієї молекули білка або однієї молекули РНК. Ці та інші функціональні молекули визначають розвиток, ріст і нормальний стан організму.
Гени людини, як і інших вищих організмів, включають Екзони і інтрони. У той час як Екзони містять кодують послідовності гена, функція нітронів залишається невідомою, а інтрони, як правило, складають основну частину гена. Нітроном були вперше виявлені в 1988 р в гені бета-глобіну миші. На кордоні екзонів і нітронів розташовується консенсусна, тобто еволюційно консервативна послідовність, яка розпізнається ферментами сплайсингу, тобто ферментами для вирізання інтронів з первинного транскрипту мРНК. На 3 "-кінців гена вже в некодирующей частині розташований сайт, що забезпечує додавання 100-200 залишків аденіну до мРНК для забезпечення її стабільності.
Для гена характерна так звана відкрита рамка зчитування, тобто наявність послідовності триплетів, що кодують амінокислоти, які не перебиває стоп-кодонами або безглуздими триплету.
Ген - ділянка молекули ДНК, що містить інформацію про первинну структуру однієї молекули білка.
5. ЗАГАЛЬНІ ВЛАСТИВОСТІ ГЕНЕТИЧНОГО МАТЕРІАЛУ. РІВНІ ОРГАНІЗАЦІЇ ГЕНЕТИЧНОГО АПАРАТУ
(ГЕННИЙ, хромосомні І геномної).
По-перше, генетичний матеріал повинен мати здатність до самовідтворення, щоб в процесі розмноження передавати спадкову інформацію, на основі якої буде здійснюватися формування нового покоління. По-друге, для забезпечення стійкості характеристик в ряду поколінь спадковий матеріал повинен зберігати постійної свою організацію. По-третє, матеріал спадковості і мінливості повинен мати здатність набувати зміни і відтворювати їх, забезпечуючи можливість історичного розвитку живої матерії в мінливих умовах. Тільки в разі відповідності зазначеним вимогам матеріальний субстрат спадковості і мінливості може забезпечити тривалість і безперервність існування живої природи та її еволюцію.
ГЕННИЙ. Елементарної функціональної одиницею генетичного апарату, що визначає можливість розвитку окремої ознаки клітини або організму даного виду, є ген (спадковий завдаток, по Г. Менделя). Передачею генів в ряду поколінь клітин або організмів досягається матеріальна спадкоємність - успадкування нащадками ознак батьків.
На хромосомний рівні організації спадковий матеріал володіє всіма характеристиками субстрату спадковості і мінливості, в тому числі і здатністю до придбання змін, які можуть передаватися новому поколінню.
Геномної РІВЕНЬ організації спадкового матеріалу, який об'єднує всю сукупність хромосомних генів, є еволюційно сформованою структурою, яка характеризується відносно більшою стабільністю, ніж генний і хромосомний рівні.
6. ГЕНЕТИЧНИЙ КОД.
Генетичний код - властивий всім живим організмам спосіб кодування амінокислотної послідовності білків за допомогою послідовності нуклеотидів.
У ДНК використовується чотири нуклеотиди - аденін (А), гуанін (G), цитозин (С), тимін (T), які в російськомовній літературі позначаються буквами А, Г, Ц і Т. Ці букви складають алфавіт генетичного коду. У РНК використовуються ті ж нуклеотиди, за винятком тиміну, який замінений схожим нуклеотидом - урацілом, який позначається буквою U (У в російськомовній літературі). У молекулах ДНК і РНК нуклеотиди шикуються в ланцюжки і, таким чином, виходять послідовності генетичних букв.
Для побудови білків в природі використовується 20 різних амінокислот. Кожен білок є ланцюжком або кілька ланцюжків амінокислот в строго певній послідовності. Ця послідовність визначає будову білка, а отже всі його біологічні властивості. Набір амінокислот також універсальний для майже всіх живих організмів.
Реалізація генетичної інформації в живих клітинах (тобто синтез білка, що кодується геном) здійснюється за допомогою двох матричних процесів: транскрипції (тобто синтезу мРНК на матриці ДНК) і трансляції генетичного коду в амінокислотну послідовність (синтез поліпептидного ланцюга на матриці іРНК). Для кодування 20 амінокислот, а також сигналу «стоп», що означає кінець білкової послідовності, достатньо трьох послідовних нуклеотидів. Набір з трьох нуклеотидів називається кодоном. Прийняті скорочення, відповідні амінокислотам і кодонам, зображені на малюнку.
Властивості ГЕНЕТИЧНОГО КОДА
1. Триплетність - значущою одиницею коду є поєднання трьох нуклеотидів (триплет, або кодон).
2. Безперервність - між триплетами немає розділових знаків, тобто інформація зчитується безперервно.
3. Неперекриваемость - один і той же нуклеотид не може входити одночасно до складу двох або більше триплетів (не дотримується для деяких перекриваються генів вірусів, мітохондрій і бактерій, які кодують кілька білків, зчитувати із зсувом рамки).
4. Однозначність (специфічність) - певний кодон відповідає тільки однієї амінокислоті (проте, кодон UGA у Euplotescrassus кодує дві амінокислоти - цистеїн і селеноцистеїн)
5. Виродженість (надмірність) - одній і тій же амінокислоті може відповідати декілька кодонів.
6. Універсальність - генетичний код працює однаково в організмах різного рівня складності - від вірусів до людини (на цьому засновані методи генної інженерії; є ряд винятків, показаний в таблиці розділу «Варіації стандартного генетичного коду» нижче).
7. Перешкодостійкість - мутації замін нуклеотидів НЕ призводять до зміни класу кодованої амінокислоти, називають консервативними; мутації замін нуклеотидів, що призводять до зміни класу кодованої амінокислоти, називають радикальними
7. ХАРАКТЕРИСТИКА гибридологический МЕТОДУ ДОСЛІДЖЕННЯ.
Гибридологический метод (метод схрещування). Розроблено Г. Менделем і є основним в генетичних дослідженнях. За допомогою схрещування можна встановити: 1) домінантою або рецессивен досліджуваний ознака (і відповідний йому ген); 2) генотип організму; 3) взаємодія генів і характер цієї взаємодії; 4) явище зчеплення генів; 5) відстань між генами; 6) зчеплення генів з підлогою.
Об'єктом дослідження Г. Мендель вибрав горох Pisumsativum, оскільки це самозапилюється рослина має багато культурних сортів, стабільно зберігають свої ознаки в потомстві, і володіє такою будовою квіток, яке дозволяє легко видаляти пильовики і наносити пилок інших сортів на рильце полукастрірованних квіток.
Протягом двох років Мендель відчував 34 сорти гороху. Переконавшись напротязі декількох циклів самозапилення в константності обраних ознак, він вибрав 22 сорти з їх контрастують парами.
У всіх схрещування Мендель проводив точний кількісний облік усіх форм другого покоління, що розрізняються за окремими ознаками: потомство від кожної рослини аналізувалося їм індивідуально, а потім підраховувалася сумарна чисельність за кожною ознакою по всій вибірці рослин.
Описуючи результати схрещувань, сам Мендель ні інтерпретував встановлені ним факти як якісь закони. Але після їх перевідкриття і підтвердження на рослинних і тваринних об'єктах, ці повторювані за певних умов явища стали називати законами спадкування ознак у гібридів.
8. законах Менделя ЗАКОНОМІРНОСТІ УСПАДКУВАННЯ за моногибридном І дигибридное схрещування.
Закон домінування - перший закон Менделя - називають також законом одноманітності гібридів першого покоління, так як у всіх особин першого покоління проявляється одна ознака. Моногібрідним називається схрещування двох організмів, що відрізняються один від одного по одній парі альтернативних (взаємовиключних) ознак. Отже, при такому схрещуванні простежуються закономірності успадкування ТІЛЬКИ двох
ознак, розвиток яких обумовлено парою алельних генів. Всі інші ознаки, властиві даними організмам, до уваги НЕ приймаються.
Закон розщеплення, або другий закон Менделя. Якщо нащадків першого покоління, однакових по досліджуваному ознакою, схрестити між собою, то в другому поколінні ознаки обох батьків з'являються в певному числовому співвідношенні: 3/4 особин матимуть домінантна ознака.
Закон незалежного комбінування, або третій закон Менделя. Вивчення Менделем однієї пари алелей дало можливість встановити ряд важливих генетичних закономірностей:
Явище домінування,
Незмінність рецесивних алелей у гібридів,
Розщеплення потомства гібридів у відношенні 3: 1 ...
А також припустити, що гамети генетично чисті, т. Е. Містять тільки один ген з алельних пари.
Однак організми розрізняються за багатьма генами. Встановити закономірності успадкування двох пар альтернативних ознак і більше можна шляхом дигибридного або полигибридного схрещування.
9. ВИЗНАЧЕННЯ: менделирующих ОЗНАКИ, ПРИКЛАДИ.
Ознаки, успадкування яких підпорядковується перерахованим закономірностям, прийнято називати
менделирующих (по імені Г. Менделя). У людини менделирующих ознаками є, наприклад, альбінізм (відсутність пігментації, що викликається рецесивним геном, колір очей, характер волосся (кучеряве або гладкі), групові відмінності по різних факторів в крові
Деякі Менделюючі ознаки людини.
1) Очі: великі (будинок), маленькі (рец);
2) Верхня повіка: нависає (будинок), нормальне (рец);
3) Вії: довгі (будинок), короткі (рец);
4) Колір очей: карі, зелені (будинок), блакитні, сірі (рец);
5) Зір: короткозорість (будинок), нормальне (рец);
6) Ніс: великий (будинок), середній, маленький (рец);
7) Вуха: широкі (будинок), вузькі (рец).
10. ГІПОТЕЗА ЧИСТОТИ ГАМЕТ.
Для пояснення закономірностей прояву і розщеплення ознак у гібридів другого покоління і в зворотних схрещуваннях Мендель запропонував гіпотезу чистоти гамет, згідно з якою, гамети кожного з батьків несуть тільки по одному з успадкованих факторів. Гібриди першого покоління (Аа) дають два типи гамет, в рівному співвідношенні містять домінантний (А) і рецесивний фактори (а).
Гібридні рослини виглядають однаково, оскільки діє закон домінування, або однаковості.
спадкові детермінанти рецесивних ознак в гібридному організмі Аа не зникають і не зливаються, а роз'єднуються з домінантними факторами в черговому циклі освіти гамет.
Для спрощення аналізу очікуваних результатів в F2 використовують так звану грати Пеннета - таблицю, перші рядки і стовпці якої відповідають різним типам жіночих і чоловічих гамет.
11. аналізує схрещування.
Аналізує схрещування - схрещування гібридної особини з особиною, гомозиготною по рецесивним аллелям, тобто "аналізатором". Сенс аналізує схрещування полягає в тому, що нащадки від аналізує схрещування обов'язково несуть один рецесивний аллель від "аналізатора", на тлі якого повинні проявитися аллели, отримані від аналізованого організму.
Для аналізує схрещування (крім випадків взаємодії генів) характерно збіг розщеплення за фенотипом з розщепленням за генотипом серед нащадків. Таким чином, аналізує схрещування дозволяє визначити генотип і співвідношення гамет різного типу, утворених аналізованої особиною.
12. Типи І ВАРІАНТИ УСПАДКУВАННЯ ОЗНАК (моногенне І полігенною, аутосомномдомінантному І аутосомноцрецессивному, зчеплених зі статтю, ГОЛАНДРІЧЕСКОЕ).
Полигенное спадкування - ознака контролюється 2 і більше генами.
Моногенне спадкування - ознака контролюється одним геном.
Типи успадкування ознак при моногенному спадкуванні:
Аутосомное успадкування. Характерні риси аутосомного успадкування ознак обумовлені тим, що відповідні гени, розташовані в аутосомах, представлені у всіх особин виду в подвійному наборі. Це означає, що будь-який організм отримує такі гени від обох батьків. Відповідно до закону чистоти гамет в ході гаметогенезу все статеві клітини отримують по одному гену з кожної алельних пари. Обгрунтуванням цього закону є розбіжність гомологічних хромосом, в яких розташовуються алельних гени, до різних полюсів клітини в анафазе I мейозу.
З огляду на те що розвиток ознаки у особини залежить в першу чергу від взаємодії алельних генів, різні його варіанти, які визначаються різними алелями відповідного гена, можуть успадковуватися по аутосомно-домінантним або аутосомно-рецесивним типом, якщо має місце домінування. Можливий також проміжний тип успадкування ознак при інших видах взаємодії алелей. При домінуванні ознаки, описаному Г. Менделем в його дослідах на горосі, нащадки від схрещування двох гомозиготних батьків, що розрізняються за домінантним і рецесивним варіантів даної ознаки, однакові і схожі на одного з них (закон одноманітності F1). Описане Менделем розщеплення за фенотипом в F2 у відношенні 3: 1 в дійсності має місце лише при повному домінуванні одного алеля над іншим, коли гетерозиготи фенотипно подібні з домінантними гомозиготами (закон розщеплення в F2) .Наследованіе рецесивного варіанти ознаки характеризується тим, що він не проявляється у гібридів F1, а в F2 проявляється у чверті нащадків.
Зчеплене з підлогою. Характер успадкування зчеплених зі статтю ознак в ряду поколінь залежить від того, в якій хромосомі знаходиться відповідний ген. У зв'язку з цим розрізняють Х-зчеплене і Y-зчеплене (голандріческое) успадкування.
Х-зчеплене успадкування. Х-хромосома присутня в каріотипі кожної особини, тому ознаки, що визначаються генами цієї хромосоми, формуються у представників як жіночого, так і чоловічої статі. Особи гомогаметний статі отримують ці гени від обох батьків і через свої гамети передають їх всім нащадкам. Представники гетерогаметного статі отримують єдину Х-хромосому від гомогаметний батька і передають її своїм гомогаметний потомству. У ссавців (в тому числі і людини) чоловіча стать отримує Х-зчеплені гени від матері і передає їх дочкам. При цьому чоловіча стать ніколи не буде наслідувати батьківського Х-зчепленого ознаки і не передає його своїм синам.
Y-зчеплене (голандріческое успадкування). Активно функціонують гени Y-хромосоми, що не мають алелей в Х-хромосомі, присутні в генотипі тільки гетерогаметного статі, причому в гемізиготність стані. Тому вони проявляються фенотипно і передаються з покоління в покоління лише у представників гетерогаметного статі. Так, у людини ознака гипертрихоза вушної раковини ( «волохаті вуха») спостерігається виключно у чоловіків і успадковується від батька до сина.
13. Класифікація ФОРМ Взаємодія генів.
Розрізняють внутріаллельное взаємодія генів (повне і неповне домінування, наддомінування, Кодомінування тд) і межаллельное взаємодія (компліментарність, епістаз і полімерія)
14. ВНУТРІАЛЛЕЛЬНОЕ ВЗАЄМОДІЯ ГЕНІВ: ПОВНЕ І НЕПОВНЕ ДОМІНУВАННЯ, Наддомінування, Кодомінування, ПРИКЛАДИ.
Повне домінування: А\u003e a; Ф: AA = АA (Горошок Менделя)
Неповне домінування: Ф: AA ≠ Aa, колір пелюсток у беладони (P-червоні і білі; F1- рожеві)
Наддомінування: A \u003e\u003e a; у Аа ознака виражений набагато сильніше у дрозофіл - АА< Кодомінування: присутність двох домінантних генів проявляється як нову ознаку, наприклад VI група крові АВ 15. плейотропії: ХАРАКТЕРИСТИКА, ПРИКЛАДИ. Синдром Марфана. Плейотропія - множинне дію гена, здатність одного спадкового чинника (гена) впливати одночасно на кілька різних ознак організму. Так, гени, що визначають забарвлення шерсті у будинкової миші, впливають на розміри тіла; ген, що впливає на пігментацію очей у млинової огнівки, має ще 10 морфологічних і фізіологічних проявів і т.д. Плейотропія часто поширюється на ознаки, що мають еволюційне значення - плодючість, тривалість життя, здатність виживати в крайніх умовах середовища. Синдром Марфана - спадкове захворювання людини, обумовлене мутацією, яка успадковується по аутосомно-домінантним типом. Характеризується надмірним зростанням пальців на руках і ногах ( «павукові пальці» або арахнодактилія). Відзначається гігантизм. Частота поширення - 1: 20000. 16. МНОЖИННІ аллель, УСПАДКУВАННЯ ГРУП КРОВІ ЗА СИСТЕМОЮ АВО. Сутність множинного алелізм. Існують приклади лише з двома аллельними станами локусу, наприклад А - а, W - w. В результаті взаємодії цих алелей спостерігалося повне або неповне домінування певної ознаки. У ряді випадків локус може мати кілька станів, т. Е. Множинний алелізм. Спадкування груп крові відбувається за типом множинного алелізм (за винятком IV групи - кодоминирование). Ген I має кілька станів: IAдомінантний, IBдомінантний, I0рецесівний Поєднання цих станів формує ту чи іншу групу крові. I0 I0 - 1 група; IА IА, IА I0 - 2 група; IВ IВ, IВ I0 - 3 група; IА IВ - 4 група (кодоминирование) 17. МЕЖАЛЛЕЛЬНОЕ ВЗАЄМОДІЯ ГЕНІВ: компліментарні, епістаз (ВИДИ Епістаз), полімери, ПРИКЛАДИ. Розвиток живої матерії на Землі відбувається в нескінченній зміні поколінь. Життя нерозривно пов'язана з розмноженням організмів. В яких би формах воно не здійснювалося, від одного покоління іншому завжди передаються загальні, характерні для даного виду ознаки і властивості. Іншими словами, потомство в тій чи іншій мірі обов'язково схоже на своїх батьків. Процес відтворення організмами в ряду послідовних поколінь схожих ознак і властивостей називається спадковістю
.
Вона проявляється у всьому тому загальному, що є між спорідненими поколіннями організмів. Таким чином, розмноження пов'язане зі спадковістю. Вже друге покоління колись виникла живої матерії було схоже на перше. Часто ознаки і властивості організмів при розмноженні відтворюються дуже стійко: діти бувають на диво схожі на своїх батьків, проте абсолютного подібності між ними ніколи не буває, завжди відрізняються один від одного з яких-небудь ознаками і діти одних і тих же батьків. Спадковість - це не просте відтворення, копіювання будь-яких незмінних властивостей і ознак організмів. Вона завжди супроводжується їх мінливістю
:
при розмноженні організмів поряд зі збереженням одних ознак змінюються інші. Не тільки відтворюється подібне, але і виникає нове. Спадковість і мінливість завжди супроводжують один одного і виявляються в процесі розмноження організмів спільно як суперечливі і в той же час нерозривно пов'язані між собою процеси. Розмноження, отже, пов'язане не тільки зі спадковістю, а й з мінливістю організмів. Наука про спадковість і мінливість організмів отримала назву генетика
(Від грец. geneticos- відноситься до походження). Головне завдання генетики
- розробка методів управління спадковістю і мінливістю для здобуття потрібних людині форм рослин, тварин і мікроорганізмів і управління індивідуальним розвитком організмів. Генетика, як будь-яка наука, має свої методи дослідження
: 1.
гибридологический аналіз, Що полягає у використанні системи схрещувань для встановлення характеру успадкування ознак і генетичних відмінностей досліджуваних організмів. Гибридологический аналіз, доповнений після робіт Г. Менделя рядом специфічних методів і прийомів дослідження спадковості, увійшов в якості найважливішої складової частини в генетичний аналіз - основний метод генетики. 2.
цитологічний метод- вивчення структур клітин в зв'язку з розмноженням організмів і передачею спадкової інформації. На основі цього методу при використанні новітніх способів вивчення хромосомних структур виникла нова наука - цитогенетика. 3.
онтогенетический методвикористовується для вивчення дії генів і прояви їх в індивідуальному розвитку організмів - онтогенезі в різних умовах зовнішнього середовища. 4.
Статистичний метод, За допомогою якого вивчають статистичні закономірності спадковості і мінливості організмів. В історії генетики можна виділити три основні періоди. Два з них, що тривали з 1900 по 1953 рік, становлять епоху класичної генетики. Третій період, що почався після 1953 р відкрив епоху молекулярної генетики. перший період(1900-1910) в розвитку генетики пов'язаний з утвердженням відкриттів Г. Менделя: принципу дискретності в передачі спадкового матеріалу і методу гибридологического аналізу. Численні досліди по гібридизації, проведені в першому десятілетііXXв. з різними рослинами і тваринами, показали, що правила в спадкуванні ознак, встановлені вперше Г. Менделем, мають універсальний характер і застосовні по відношенню до всіх організмів, які розмножуються статевим шляхом. Отже, закони спадковості єдині для всього органічного світу. У цей період закономірності успадкування ознак вивчають на рівні цілісного організму і не пов'язують з будь-якими матеріальними структурами клітини. Передачу і поширення в поколіннях спадкових факторів розраховують за допомогою буквених схем і формул. Найважливіше значення для подальшого розвитку генетики мала висунута в цей час (1901-1903) голландським вченим Гуго Де-Фріз теорія мутацій. Данська генетик В. Иоганнсен на основі своїх дослідів з вивчення успадкування ознак в популяціях і чистих лініях квасолі (1903) розробив і запровадив в генетику поняття ген,
генотип,
фенотип
(1909). другий
період
(1911-1953) пов'язаний з встановленням матеріальних основ спадковості. Ще в перше десятиліття розвитку генетики (1902-1907) Т. Бовери, У. Сеттон і Е. Вільсон обгрунтували хромосомну теорію спадковості. Було з'ясовано, що між поведінкою спадкових факторів і хромосом в процесах клітинного ділення (мітоз) і утворенням статевих клітин (мейоз), що передаються наступним поколінням, існує певний зв'язок. Для вивчення явищ спадковості в цей час стали користуватися цитологическими методами. Відбулося об'єднання методу генетичного аналізу з цитологічним методом. Так в генетиці виникло цитогенетическое напрямок. Було встановлено, що спадкові чинники знаходяться в клітці. Вивчення спадковості піднялося на більш високий рівень. Вирішальне значення для обґрунтування і затвердження хромосомної теорії спадковості мали почалися в 1910 р дослідження американського генетика Т. Моргана з плодовою мушкою дрозофіли. У роботах Т. Моргана і його учнів поняття спадкового чинника (гена) отримало матеріальне втілення. Хромосомна теорія спадковості встановила, що гени знаходяться в хромосомах і розташовані в них в лінійному порядку. Гени утворюють стільки груп зчеплення, скільки пар гомологічних хромосом є у даного виду. Гени, що знаходяться в одній групі зчеплення, завдяки явищу перехрещення (кросинговеру) можуть рекомбінуватися; величина рекомбінації - функція відстані між генами. До початку 20-х рр. у дрозофіли було виявлено і локалізовано у всіх чотирьох групах зчеплення кілька сотень генів. Встановлені на плодової мушки принципи визначення місця розташування генів в хромосомах були перенесені на інших тварин і рослинні об'єкти і виявилися вірними для всіх видів організмів. третій
період
в розвитку генетики, що почався після 1953 р пов'язаний з використанням методів і принципів досліджень точних наук: хімії, фізики, математики, кібернетики і т. д. Стали широко застосовувати електронну мікроскопію, рентгеноструктурний аналіз, швидкісне центрифугування, метод радіоактивних ізотопів, чисті препарати вітамінів, ферментів і амінокислот і т. д. Аналіз матеріальних основ спадковості перейшов на молекулярний рівень вивчення структурної організації живої матерії. Розвиток сучасної генетики характеризується проникненням молекулярних принципів досліджень в усі області вчення про спадковість. Широкий розвиток отримали дослідження з таких проблем, як штучний синтез гена поза організмом, продовжений мутагенезу, гібридизація соматичних клітин, отримання гаплоїдних рослин при культивуванні пиляків, механізми регуляції активності і дію генів в процесах індивідуального розвитку, молекулярні основи рекомбінації, репарацій (відновлення) первинних ушкоджень генетичного матеріалу, генна інженерія, штучний синтез нуклеїнових кислот і білків і ін. Для сучасного стану генетики характерні, з одного боку, вплив на неї принципів і методів досліджень точних наук, а також все зростаюча зв'язок її з іншими біологічними науками; з іншого боку, в самій генетиці йде надзвичайно швидкий процес диференціації і перетворення окремих розділів і напрямків досліджень в самостійні науки. Так, в короткий термін поряд із загальною генетикою, генетикою тварин і генетикою рослин виникли цитогенетика, генетика людини, медична генетика, космічна генетика, генетика популяцій, еволюційна генетика, біохімічна генетика, генетика мікроорганізмів, генетика вірусів, генетика соматичних клітин, генетика фотосинтезу, екологічна генетика, математична генетика, генетика поведінки і т. д.