Kada se monohibrid prelaze homozigotni pojedinci. Monohybrid cross
Prilikom ukrštanja bijelih andaluzijskih pilića s crnim pijetlovima, dobivene su kokoške sa sivim perjem. Uzgajanjem sivih pilića dobijaju se potomci. F 2. 1) Koji je genotip sivih pilića? 2) Koliko se klasa genotipova i fenotipa formira u F 2? 3) Šta su pilići F 2 će dati ne-deljenje potomaka?
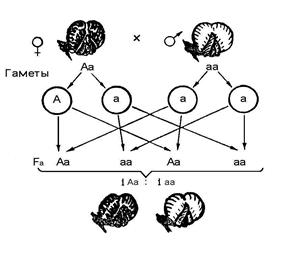
Nasleđivanje perja u andaluzijskim pilićima je prikazano na sl. 2. Linija rasuđivanja i određivanje uzoraka nasleđivanja karaktera slični su zadatku 2. Budući da uslovi problema impliciraju da su heterozigoti srednji u boji, genotip roditeljskih formi odgovara formulama aa i AA. Prelazak belih i crnih pilića daje sivo potomstvo F 1 genotip Aa. Heterozigotni pilići formiraju dvije vrste gameta. Kao rezultat toga F 2 formirao 3 klase genotipova u odnosu 1AA: 2AA: 1AA. Za razliku od prethodnog slučaja, fenotip homozigotnih oblika razlikuje se od fenotipa heterozigotnih jedinki, a bijeli, sivi i crni pilići pojavljuju se u potomstvu u omjeru 1: 2: 1. ne potpuna dominacija Cijepanje genotipa odgovara cijepanju fenotipa. Slika takođe pokazuje da je u F 3bijeli i crni pilići proizvode piliće sličnog tipa, a kada prelaze heterozigotne piliće, fenotipsko cijepanje se opet vidi u omjeru 1: 2: 1.
Odgovori:1) sivi pilići imaju genotip Aa; 2) u F 2 Formiraju se 3 klase genotipova i 3 klase fenotipa; 3) crne i bele piliće neće biti podijeljene.
Koristeći primjer ova dva zadatka, možemo izvući zaključke:
1. Prilikom prelaska homozigotnih biljaka (ili životinja) i njihovih hibrida među sobom, nasleđe osobina slijedi Mendelove zakone.
2. Fenotip hibrida F 1 ukazuje na pravac i stepen dominacije gena (osobina).
3. Vydellenie forme sa recesivnom osobinom moguće je samo u potomcima heterozigotnih oblika, dok homozigoti daju stabilno potomstvo
Sl. 11Nasleđivanje perja u andaluzijskim pilićima
prema vrsti nepotpune dominacije
.Zadatak 3. Analiziranje prelaza.
U analizi ukrštanja biljaka graška sa ljubičastim cvetovima, dobijeno je 18 biljaka sa ljubičastim i 18 biljaka sa belim cvetovima. 1) Koji je genotip analizirane biljke? 2) Koliko tipova gameta ima ova biljka? 3) Koji deo biljaka dobijenih analizom krstova će dati neljubazno potomstvo?
U ovom zadatku, neophodno je odrediti genotip roditeljskih oblika zbog poznavanja karakteristika analizirajućeg križa. Budući da je po definiciji analizirajući križ hibridizacija s homozigotnom recesivnom formom, genotip paternalnog oblika je aakoji odgovara fenotipu bijelih cvjetova. Da bi se odredio genotip purpurno-cvetnog oblika majke, potrebno je zapamtiti koji su rezultati dobiveni analizom križanja s dominantnim homozigotnim i heterozigotnim oblikom:
R ♀ AA x ♂ aa P ♀ Aax ♂ aa
A 1 Aa A 1 Aa Aa
Kao što se može vidjeti iz dijagrama, ako je biljka s dominantnim fenotipom homozigotna (AA), svi hibridi imaju dominantnu osobinu; i kada se ukrsti sa heterozigotnim oblikom u potomstvu, razdvajanje se posmatra u odnosu 1: 1.
Prema stanju problema, kao rezultat analize križanja, u potomstvu je uočeno 18 ljubičastih i 18 belih cvetnih oblika, što odgovara podeli 1: 1 i ukazuje na heterozigotnost materinskog oblika. Dalje, tok zadatka je ilustrovan na sl. 3. Analizirani oblik formira dvije vrste gameta. Pri spajanju gameta nastaju heterozigotne ljubičaste cvjetnice i homozigotne bijele cvjetnice. Kao što je pokazano u prethodnim primjerima, samo homozigotne biljke proizvode stabilno potomstvo, dok heterozigotne pokazuju rascjep u narednim generacijama. Stoga će nekodificirano potomstvo dati samo 18 biljaka sa bijelim cvjetovima.
Sl. 12 Hibrid sa križanjem F 1 sa recesivnim
roditeljski oblik (analizirajući križ)
Odgovori: 1) analizirana biljka ima genotip Aa; 2) analizirana biljka formira dvije vrste gameta; 3) kao rezultat analize križanja 1/2 potomstva, tj. 18 biljaka će biti homozigotne i dati će potomstvo koje nije cepanje.
Zadatak 4. Hibridni prelaz.Prilikom prelaska raznih paradajza sa crvenim okruglim plodovima sa biljkama žutog kruškolikog voća, dobijeni su crveno plodni hibridi sa okruglim plodovima. Kao rezultat samooprašivanja dobijeno je 80 hibrida. F 2. 1) Koji dio postrojenja F 1 bio digerozigot? 2) Koliko vrsta gameta formiraju biljke? F 1? 3) Koliko klasa genotipova formira hibride F 2? 4) Koje je fenotipsko cepanje uočeno u F 2? 5) Koliko biljaka F 2 će imati crveno voće u obliku kruške?
Da bi se riješio ovaj problem, potrebno je odrediti genotipove roditeljskih oblika. Označite gene koji određuju znak boje plodova slova A, a oblik ploda - pismo U. Smjer i stupanj dominacije će se odrediti fenotipom hibrida F 1. Kako su hibridi prve generacije pokazivali znakove crvene boje i okruglog oblika, ovi znakovi i odgovarajući geni pokazuju potpunu dominaciju. Označavamo gene paradajza na sledeći način:
A - crveno bojenje voća, a - žuta boja ploda;
U - okruglog oblika, u - oblik kruške.
Crossbred biljke su diploidne, stoga definiramo genotip majčinog oblika sa crvenim okruglim plodovima kao AABBi očinski sa žutim kruškolikim voćem - aavv. U ovom primjeru, znakovi su naslijeđeni nezavisno i distribucija gena je povezana sa nezavisnom divergencijom dva para homolognih hromozoma u profazi mejoze. Za bolje razumijevanje materijala preporučuje se da alelni geni lokalizirani u različitim parovima kromosoma budu naglašeni različitim linijama koje simboliziraju pripadnost različitim homolognim parovima (Slika 5). Kada se gamete formiraju u mejozi, jedan hromozom iz svakog para homolognih hromozoma i jedan alelni gen, nasumce se distribuira svakom gametu. Homozygous roditeljski oblici čine jedan tip gameta. Kada se stapaju, formiraju se biljke koje su heterozigotne za dva para gena - diheterozigot. Biljke F 1 jednaka za svaki par znakova, tj. manifestovani
I Mendelov zakon.
Digesterozigotne biljke F 1 oblik 2 2 = 4 vrste gameta, dok je kombinacija hromozoma i gena različitih parova u gametima slučajna i jednako vjerojatna. Odrediti genotipove i fenotipove hibrida F 2 Neophodno je izgraditi mrežu Penneta. Rešetka se sastoji od 16 (4 2) ćelija, jednako je vjerovatan i izgled odgovarajućeg genotipa. Brojanje pokazuje da se 9 (3 2) genotipova formira tokom dihibridnog križanja. Prema fenotipu, potomstvo je podijeljeno u 4 klase u omjeru:
Biljke sa crvenim krušnatim plodovima imaju genotip A-cc i čine 3/16 potomke, tj. 15 biljaka.
Odgovori1) sve biljke F 1 diheterozygous; 2) hibride prve generacije formiraju 4 tipa gameta; 3) u F 2 9 genotipova i 4 klase fenotipova formiraju se u omjeru 9: 3: 3: 1; 5) u F 2 Pojavit će se 15 biljaka sa crvenim kruškolikim plodovima.

Sl. 5 Nasleđivanje znakova boje i oblika voća u paradajzu
Eksperimenti Mendel je potrošio na grašak. Prilikom prelaska sorti graška sa žutim i zelenim sjemenkama (homozigotni organizmi ili čiste linije prekrižene), svi potomci (tj. Hibridi prve generacije) su se pokazali kao žuti sjemeni. Čini se da suprotni znak (zeleno sjeme) nestaje. Otkriveni uzorak je dobio ime pravilo uniformnosti (dominacije) hibrida prve generacije (iliprvi zakon G.Mendela ).
Eksperimenti na križanju su zabilježeni u obliku dijagrama:
A - žuta boja gena
a - gen zeleni
R - (roditelji - roditelji)
F - (filii - djeca)
P AAA x aaa
F 1 AA - 100% žuta
Dakle, svi hibridi prve generacije su homogeni: heterozigotni u svom genotipu i dominantni u svom fenotipu.
Tako se prvo pravilo (zakon) Mendelove uniformnosti hibrida prve generacije može formulisati na sledeći način: kod prelaska homozigotnih pojedinaca koji se međusobno razlikuju za jedan par alternativni znakovi, svi potomci u prvoj generaciji su uniformni u oba fenotipa i genotipa
Pravilo razdvajanja Drugi zakon Mendela
Ako pređete hibride prve generacije, u drugoj generaciji, pojavljuju se pojedinci, i sa dominantnim i recesivnim osobinama, tj. postoji razdvajanje u određenom numeričkom odnosu. U eksperimentima sa zrnom graška žuto seme je tri puta više od zelenog. Ovaj obrazac se naziva drugi zakon (pravilo) Mendela, ili zakon (pravilo) razdvajanja.
R a Aa x. Aa
G (A) (a) (A) (a)
F 2 AA; Aa, Aa; aa
žuto zelena
Cepanje fenotipa 3: 1, genotip 1AA: 2Aa: 1aa
Drugi zakon (pravilo) Mendela : Kada su dva heterozigotna jedinke ukrštena, analizirana jednim alternativnim parom znakova (tj. Hibridi), očekuje se da će potomstvo biti razdvojeno fenotipom 3: 1 i genotipom 1: 2: 1.
Di-i polybreeding. Treći zakon Mendela
U dihibridnom prelazu, roditeljski organizmi se analiziraju pomoću dva para alternativnih osobina. Mendel je proučavao takve znakove kao što su bojenje sjemena i njihov oblik. Prilikom ukrštanja graška sa žutim i glatkim sjemenkama sa graškom sa zelenim i naboranim sjemenkama, u prvoj generaciji svi potomci su bili homogeni, dominantne osobine – žuta boja i glatke forme. Prema tome, kao i kod monohibridnog križanja, pravilo jedinstvenosti hibrida prve generacije ili pravilo dominacije.
Žuti gen
a - gen zeleni
B - gen glatkog oblika
c - gen naboranog oblika
R JAAVV x Đavav
g. Ch. h bora
G (AB) (AB)
F 1 AaB - glatka žuta
Kod križanja hibrida prve generacije, razdvajanje fenotipa se dogodilo između sebe:
R В AaVv x AaVv
9 komada - glatka žuta
3 komada - žuta naborana
3 komada - zelena glatka
1 dio - zelena naborana
Iz ovog prelaza može se vidjeti da u drugoj generaciji nije bilo samo pojedinaca sa kombinacijom roditeljskih osobina, već i pojedinaca s novim kombinacijama osobina.
Pored toga, Mendel je otkrio da je svaki par osobina (boja i oblik) dao podjelu približno u omjeru 3: 1, to jest, kao kod monohibridnih križanja. Iz ovoga je zaključeno da se svaki par alternativnih osobina nasleđuje nezavisno jedan od drugog tokom di- i polybreeding.
Treće pravilo ili treći zakon Mendela je formulisan na sledeći način: kada se prelaze homozigotni pojedinci koji se razlikuju u dva (ili više) parova alternativnih osobina, u drugoj generaciji se posmatra nezavisno nasleđivanje i kombinacija osobina ako su geni koji ih određuju locirani u različitim homolognim hromozomima.
Pored zakona, Mendel je formulisao dve hipoteze: faktorijsku hipotezu i "čistoću gameta", uz pomoć kojih je pokušao da objasni ustanovljene zakonitosti.
Faktorska hipoteza pokazuje da ćelije sadrže faktor (gen) koji nosi osobinu. Roditelji prenose potomcima ne znake, već ove faktore.
Hipoteza "čistoća gameta"» : organizam na svakoj osobini nosi dva nasljedna faktora (jedan od oca, drugi od majke). Ovi nasledni faktori, dok su u ćelijama, ne spajaju se jedan sa drugim i, pri formiranju gameta, divergiraju se u različite gamete.
Analyzing Crosses
Recesivni alel se manifestuje samo u homozigotnom stanju. Dakle, o genotipu izlaganja tela recesivno svojstvo može se suditi po fenotipu.
Homozigotni i heterozigotni pojedinci koji pokazuju fenotipske dominantne osobine ne razlikuju se. Da bi se odredio genotip izvršena je analiza križanja i genotip roditelja određen potomstvom.
Analizirajući križanje, pojedinac čiji genotip nije jasan, ali mora biti razjašnjen, prelazi se recesivnom formom. Ako su svi potomci homogeni od takvog krsta, onda je analizirana osoba homozigotna, ako se pojavi razdvajanje, onda je heterozigotna
P x AA x ♂ aa
P a Aa x a aa
G (A) (a) (a)
Kao što se može vidjeti iz dijagrama, podjela 1: 1 karakteristična je za potomstvo heterozigotnog pojedinca pri analizi križa.
Hromozomska teorija nasljednosti
Plan
1. Sex kao nasljedna osobina.
2. Određivanje pola.
3. Nasleđivanje znakova vezanih za pod i ograničeno na pod.
4. Spajanje gena. Eksperimenti i vladavina Morgana.
5. Glavne odredbe kromosomske teorije nasljednosti.
Kromosomska teorija nasljednosti je dio genetike o lokalizaciji naslijeđenih faktora u kromosomima stanica.
Njegova suština leži u činjenici da su svi znaci i svojstva živih organizama linearno određeni genima koji se nalaze u hromozomima ćelije. Njen osnivač je bio američki genetičar Tomas Morgan, teorija koju je razvilo njegovo osoblje početkom dvadesetog veka.
Rod kao nasljedna osobina
Jedan od znakova kod mnogih živih organizama je rod (muški i ženski).
Rod je kombinacija morfoloških, fizioloških, biohemijskih i ponašajnih znakova organizma koji uzrokuju reprodukciju (reprodukciju).
Rod je genetski determinisana osobina, tj. pol embriona određuje genetski mehanizam - kombinacija seksualnih hromozoma. Polni hromozomi određuju primarne polne karakteristike.
Pod njihovim uticajem formiraju se polne žlijezde (testisi i jajnici), koje izlučuju muške i ženske spolne hormone. Polni hormoni određuju razvoj sekundarnih spolnih karakteristika jednog ili drugog pola i manifestuju se u organizmu tokom puberteta (kod ljudi u dobi od 12 do 15 godina). Sekundarne seksualne karakteristike - razlika u veličini i proporcijama tela, ton glasa, stepen razvoja kose, itd.
Određivanje pola
U većini organizama, spol je određen u vrijeme oplodnje (singamno) i reguliran je kromosomskim skupom zigota, to se naziva kromosomski tip određivanja spola.
Kod ljudi i sisara, ženke u kariotipu imaju dva X - kromosoma (XX), a muški X i Y - hromozomi. Ženke su homogametične jer formiraju samo jednu vrstu gameta, a mužjaci su heterogametni, jer formiraju dvije vrste gameta.
Kod ptica i leptira heterogametni seks je ženski, a homogametni spol je muški. Oni imaju seksualne hromozome označene kao Z i W.
Kod nekih buba, kukaca, ženke imaju sastav polnih hromozoma dvadesetog, ali mužjaci nose samo jedan od njih - (XO).
Veoma poseban tip određivanja pola se nalazi kod divljih ptica, posebno u pčelama. Kod ovih insekata, ženke se razvijaju od oplođenih jaja, a njihove ćelije su diploidne, a mužjaci se razvijaju iz partnenogenetskih jaja i imaju haploidne ćelije.
Kromosomska teorija rodnog nasleđivanja tvrdi da su geni koji određuju razvoj pola lokalizovani u spolnim hromozomima. Kod ljudi, geni odgovorni za razvoj ženskog pola su u X hromozomu, a muški geni u Y kromosomu.
Glavni tipovi kromosomskog određivanja pola
Situacija je drugačija u Drosophili. Kod proučavanja nasleđivanja pola u mušicama Drosophila, utvrđeno je da ženka ima dva identična X hromozoma, a muški X i Y hromozomi. U Drosophili, geni koji određuju ženski pol su u X hromozomu, a Y genom je genetski inertan, a geni koji određuju razvoj muškog pola su u autosomima. Oplodena jajašca sa diploidnim setom hromozoma i dva X-hromozoma daju ženke (2A + XX), a imaju diploidni set autosoma i jedan X-hromozom daju mužjacima (2A + XY).
Eksperimenti su potvrdili da što je više skupova autosoma, više je izraženih muških znakova. Iz ovoga je zaključeno da Y - hromozom u muvi - Drosophila nije bitan za određivanje muškog pola.
Pored sinamičke metode za određivanje pola, postoji i epigamička metoda za razlikovanje spola, koja je određena utjecajem uvjeta okoline, na primjer, u morskoj lišajskoj jagodici Bonnelia je određena uvjetima za razvoj jaja (progamska metoda, kao kod rotifera).
Nasleđivanje seksualno povezanih osobina
Vezani za pod nazivaju se znaci, čiji je razvoj posljedica gena lociranih u spolnim hromozomima.
Ako se gen nalazi u Y hromozomu, onda je nasleđen kod ljudi, kod sisara od oca do sina, a kod ptica od majke do kćeri.
Ako se gen nalazi na X hromozomu, nasleđivanje će biti teže. Po prvi put, karakteristike nasleđivanja gena povezanih sa X hromozomom otkrivene su u eksperimentima na Drosophili prilikom proučavanja nasleđa boje očiju. Crvena boja očiju dominira nad belom bojom.
Prilikom križanja homozigotne crvenooke ženke sa mužjakom bijele očiju u F1, svi potomci su crvenooki.
R А H A H ♂ HªU
G (X A) (Xª) (Y)
F 1 X A X x x A Y
G (X A) (Xª) (X A) (Y)
F 2 H A H, H A, H A U, HªU
cr. cr. cr. bijela
Kada se pojavi hibridizacija hibrida prve generacije u F2, javlja se fenotipska podjela: 3 crvenooke osobe: 1 bijelo-eyed, a samo muškarci su bijelo-eyed.
Kada je recipročno (obrnuto) ukrštanje, kada je ženka homozigotno za belooki gen, križana sa crvenookim muškim, podela u F1 fenotipom je 1: 1. U ovim belokim će biti samo muškarci. Kada se prelaze muve F1 jedna sa drugom u F2, biće mušice sa oba znaka u jednakom odnosu 1: 1, kako među ženama tako i među muškarcima.
R ª HªHª x ♂H A U
G (Xª) (X A) (Y)
F 1 H A Hª h
G (X A) (Xª) (Xª) (Y)
F 2 H A, HªHª, H A U, HªU
cr. bijela cr. bijela
Iz rezultata Morganovog eksperimenta možemo zaključiti da su geni za bojenje očiju smješteni samo na kromosomu spola X. \\ t Rezultati križanja zavise od toga koji rod nosi dominantnu osobinu.
Rodno povezano nasljeđivanje mora se razlikovati od rodno ograničenog nasljeđivanja.
Razvoj osobina (ograničen po polu) je zbog gena koji se nalaze u autosomima (a ne u spolnim hromozomima), ali manifestacija osobina u fenotipu je pod jakim uticajem roda, tj. Razvoj osobina zavisi od uticaja polnih hormona.
Na primjer, ćelavost od čela, tipična za muškarce. Kod muškaraca, pod utjecajem muških spolnih hormona (testosteron), gen se ponaša kao dominantan. Drugi primjer, ton glasa - bariton i bas - tipičan je samo za muškarce.
Gene coupling. Eksperimenti i vladavina Morgana
Studija o nasleđivanju povezanom sa spolom stimulisala je proučavanje veze između gena koji se nalaze u autosomima.
Za svaki organizam karakteristična je postojanost kromosoma u kariotipu. Geni koji određuju znakove organizama mnogo su veći od kromosoma. Na primer, Drosophila fly ima 8 hromozoma u somatskim ćelijama i oko 1000 gena, tako da u svakom hromozomu ima mnogo gena.
Geni lokalizovani u istoj hromozomskoj formi povezujuće grupe.
Broj grupa kvačila je jednak haploidnom broju hromozoma.
Nasleđivanje gena lociranih u jednom hromozomu naziva se povezano nasljeđivanje.
Početkom dvadesetog veka, T. Morgan i njegovi saradnici opisali su fenomen povezivanja gena - zajednički transfer genskih grupa iz generacije u generaciju.
Eksperimenti su izvedeni na mušicama voćnih mušica, uzimajući u obzir dva para alternativnih osobina:
Boja karoserije - siva (B) i crna (u)
Dužina krila - normalna (V) i kratka (v)
U muhu - Drosophila, boja tijela i dužina krila su u jednom paru homolognih kromosoma, tj. pripadaju istoj grupi kvačila, što je dokazano u eksperimentima. Evidencija genotipova sa kvačilom modifikovana: genotip je snimljen BV
(dvije crtice ukazuju da je organizam diploidan).
Prilikom prelaska leti siva boja tijela i normalna krila s crnim mušicama i kratkim krilima u F1, sve mušice su imale sivu boju tijela i normalna krila.
Snimanje uzorka ukrštanja:
ser.nor cher
G ( Bv) (bv)
Proveo je analizu krstova. U prvom slučaju, hibridni mužjak sa sivim tijelom i normalnim krilima je prešao
sa crnom ženkom sa kratkim krilima, i kao rezultat križanja, dobijene su dvije vrste potomaka, slično roditeljima u omjeru 1: 1.
Sivi gen tela i gen normalne strukture krila prenose se zajedno, a crna boja tela i gen kratkih krila su takođe zajedno. Ovo iskustvo pokazuje full grip. Razlog za to leži u činjenici da geni odgovorni za dva različita svojstva leže na istom hromozomu. To se može vidjeti u cijeloj shemi spajanja:
G ( bv) ( Bv) (bv)
Drugi rezultat se dobija ako za analizirajući križ uzmemo hibridnu ženku sa sivim telom i normalnim krilima i recesivnim mužjakom na oba osnova.
U ovom slučaju postoje četiri tipa potomaka u omjeru:
siva krilca - 41, 5%; siva kratka krila - 8, 5%,
crno dugokrvni - 8, 5%, crno krilati - 41, 5%.
U ovom slučaju dolazi do nepotpunog povezivanja:
P VBV ♂ bv
G ( Bv) (bv)
necrosses. (\\ T bv)
(Bv) (bV)
F 1 ═ ═ ═
41,5% 41,5% 8,5% 8,5%
Razlog nepotpune povezanosti gena je prelazak (hromozomski prijelaz) i razmjena područja između homolognih kromosoma u podjeli mejoze profaze I kada zrele ćelije zametka. Prelazak kod ženki, kod muškog prelaza Drosophila se ne dešava (utvrđeno je da je manja učestalost križanja karakteristična za pojedince heterogametnog pola).
Kod proučavanja rezultata križanja, dobijeni su 4 fenotipa među potomcima:
17% (8, 5 + 8, 5) pojedinaca je formirano od crossover gamete
83% (41, 4 + 41, 5) jedinki je formirano od ne-makroskopskih gamete.
Učestalost križanja zavisi od udaljenosti između gena u hromozomu. Udaljenost između gena se izražava kao procenat prelaza između njih i označava se u morganima.
Morganova studija povezanosti gena je prirodni biološki fenomen.
Geni lokalizovani u jednom hromozomu su nasleđeni, povezani, a sila spajanja zavisi od udaljenosti između gena. Ovaj uzorak se zove Morganovo pravilo.
Glavne odredbe kromosomske teorije nasljednosti
Glavne odredbe hromozomske teorije nasleđa su sledeće:
Nosioci nasljednih informacija su hrom
soma i geni koji se nalaze u njima,
Geni se nalaze u hromozomu u linearnom redosledu
prijatelj u određenim lokusima. Alelni geni zauzimaju
identični lokusi homolognih hromozoma,
Geni koji se nalaze u istoj grupi hromozoma
spojiti i naslijediti uglavnom zajedno. Number of
adhezijske grupe su jednake haploidnom skupu kromosoma,
Moguća je razmena homolognih hromozoma.
kami - prelazi preko, što prekida vezu gena.
Snaga zavisi od udaljenosti između gena,
Procenat prelaza je proporcionalan razmaku između
geni. Za jedinicu rastojanje uzima se kao 1 morganid,
koji je jednak 1% crossover,
U slučaju nepotpune adhezije u sumi, vjerovatnoća nekroze
gamete (hibridi) su uvijek veće od 50%,
Na udaljenosti od 50 morganida i više, znakovi nasljeđuju
nezavisno, uprkos tome što je lokalizovan u 1 hromo-
Molekularna genetika
Plan
1. Faze razvoja molekularne genetike.
2. Genetski kod i njegova svojstva.
3. Funkcionalna genetska klasifikacija gena.
4. Funkcionalna genska aktivnost (ekspresija gena).
5. Regulacija ekspresije gena u pro - i eukariota.
Molekularna genetika se bavi proučavanjem strukturalne i funkcionalne organizacije genetičkog aparata ćelija i mehanizma za implementaciju nasljednih informacija.
Faze razvoja molekularne genetike
Molekularna genetika nastala je iz biohemije i formirana je kao nezavisna nauka 50-tih godina prošlog veka. Rođenje ove nauke povezano je sa nizom važnih bioloških otkrića:
1). U 20-40 godina. U dvadesetom veku, utvrđeno je da je nosilac nasledne informacije u ćeliji molekul DNK, a ne protein, kao što se ranije mislilo. Dobiveni su direktni dokazi o ulozi DNK u nasleđu. To su fenomeni transformacije, transdukcije, seksualnog procesa u bakterijama, strukture virusa i otkrića gotovo potpunog identiteta hemijskog sastava DNK (ali ne proteina) u svim članovima date biološke vrste.
Podaci o tipskoj specifičnosti strukture DNK dobiveni su početkom pedesetih godina prošlog stoljeća Erwinom Chargaffom i njegovim suradnicima. Bilo koja DNK se sastoji od četiri tipa nukleotida koji sadrže jednu od četiri azotne baze: adenin, timin, gvanin, citozin. Chargaff i saradnici su otkrili da su odnosi između nukleotida koji čine DNK isti unutar vrste i variraju između vrsta. Pored toga, broj adenilnih nukleotida jednak je broju timidila, a broj guanila jednak je broju citozila. Ovo otkriće je bio glavni ključ za identifikaciju strukture molekula DNK.
Transformacija kod bakterija to je prenos nasljednog materijala iz jedne bakterijske ćelije u drugu. Godine 1928. Fred Griffith je proučavao transformaciju dva soja pneumokoka: virulentan (uzrokuje smrt miševa) i avirulentan (ne izazivajući smrt).
Ako ubijete virulentne bakterije i ubrizgate ih zajedno sa živom avirulentnom bakterijom, onda će neke od životinja i dalje umirati. Ovo iskustvo je pokazalo da je dio genetskog materijala iz virulentnih bakterija prodro u avirulentne bakterije i učinio ih virulentnim. Nakon što su virulentne ćelije podeljene na odvojene komponente: ugljene hidrate, lipide, proteine i DNK, otkriveno je da DNK daje živim bakterijama genetsku osobinu koju ranije nisu imali.
Dakle, kada se virusi koji se množe u ćelijama mobilnih kultura unose u nepokretne bakterije, neke od bakterija postaju pokretljive.
Seksualni proces u bakterijama - konjugacija, kada postoji razmjena molekula DNK, i iako nove bakterije nisu formirane, njihov nasljedni materijal se mijenja, jer dolazi do rekombinacije genetskog materijala.
Struktura virusa ili bakteriofaga: virus se sastoji od DNK molekule (ili RNK), zatvorene u proteinskoj ljusci. Uz pomoć radioizotopa, dokazano je da je DNK koja prodire u ćeliju, dok proteinska membrana ostaje vani. Kao rezultat, formiraju se nove fagne čestice. Tako je DNK nosilac genetske informacije i stvara nove bakteriofage.
Indirektni dokazi: somatske ćelije različitih organa sadrže istu količinu DNK, dvostruko više nego u spolu. Broj proteina varira i nije uvijek veći nego u genitalnom.
Sljedeća faza u razvoju molekularne genetike povezana je s tako važnim otkrićem kao što je uspostava strukturne organizacije DNA molekula 1953. godine. Crick i Watson su otkrili da se DNK sastoji od dva spiralno uvijena lanca.
Trenutno je uobičajeno govoriti o primarnoj, sekundarnoj i tercijarnoj strukturi DNK: Primarna struktura DNK je linearni polinukleotidni lanac u kojem su mononukleotidi povezani 3 ', 5' -fosfoeter vezama. Crickov i Watsonov model je sekundarna struktura DNK koja se zasniva na principu komplementarnosti i zove se dvostruka spirala. Jedan zavoj spirale sastoji se od 10 nukleotida, veličina zavojnice je 3.4 nm.
Dodatno prostorno strukturiranje dvolančane molekule DNA (u kondenzovanim hromozomima) da bi se formirao superkoil je tercijarna struktura.
3) Određivanje pravca prenosa informacija - “jedan gen - jedan enzim” - Beadle i Tatum - 50s. (Trenutno jedan gen - jedan polipeptid).
Dekodiranje genetskog koda - Nirenberg, Ochoa (Do 1964. godine dekodirane su šifre za sve aminokiseline). Elucidacija mehanizma ekspresije gena u prokariotima i njegovoj regulaciji - Francois Jacob i Jean Monod - 50s.
5) .70-te godine i do danas - identifikacija karakteristika ekspresije gena kod eukariota. Razvoj genetskog inženjeringa.
Genetski kod i njegova svojstva
Genetski kod je sistem za snimanje informacija o sekvenci rasporeda aminokiselina u proteinima koristeći sekvencu rasporeda nukleotida u molekuli DNK.
Svojstva genetskog koda :
1. Triplet koda - svaka aminokiselina je kodirana nizom od tri nukleotida, nazvanim triplet ili kodon.
2. Kod je degeneriran - svaka aminokiselina je kodirana više od jednog kodona. (Izuzetak su aminokiseline metionin i triptofan. Oni su kodirani jednim kodonom.)
3. Kod je univerzalan - iste trojke kodiraju iste aminokiseline u svim organizmima.
4. Genetski kod se ne preklapa - trojke nukleotida se ne preklapaju. Neki virusi su se preklapali, ali ovo je izuzetak.
5. Postoje startni i terminalni kodoni. Početak - jedan za sve - AUG. Terminal tri vrste - UAG, UAA, UGA.
Funkcionalna genetska klasifikacija gena
Trenutno se gen smatra jedinicom funkcioniranja nasljednog materijala. Gen je deo molekula DNK koji je odgovoran za sintezu jednog polipeptida.
Postoje tri vrste gena:
Structural;
Modulatori;
Regulatori.
Strukturni geni nose informacije o aminokiselinama u proteinima, enzimima, kao i sekvenci nukleotida u molekulima r-RNK i t-RNA.
Modulatorni geni utiču na funkcionisanje strukturnih gena, oni mogu da pomeraju razvojni proces osobine u jednom ili drugom pravcu. Podijeljeni su na:
1) inhibitori ili supresori (epistatski geni),
2) pojačivači - mogu povećati sposobnost strukturnih gena za mutacije,
3) modifikatori - mogu uticati na strukturne gene tipa komplementarnosti.
Regulatorni geni kontrolišu sintezu regulatornih proteina, kao i inkorporaciju različitih strukturnih gena u proces individualnog razvoja.
Svojstva gena:
1. Diskretnost djelovanja - svaki gen djeluje kao nezavisna jedinica nasljednosti.
2. Stabilnost - u odsustvu mutacija, gen se prenosi u nizu generacija u nepromijenjenom obliku.
3. Specifičnost akcije - svaki gen utiče na razvoj određene osobine.
4. Pleiotropija - sposobnost jednog gena da istovremeno obezbedi razvoj nekoliko znakova (Marfanov sindrom).
5. Prisustvo u obliku dva u diploidnom i većeg broja alela u poliploidnim organizmima.
6. Dozira se efekat gena, kada se mijenja broj doza gena u tijelu, promjena se mijenja (na primjer, do tri doze gena u kromosomu 21 u porastu Downe bolesti).
Funkcionalna genska aktivnost ili ekspresija gena
U prokariotima se provodi u dvije faze: transkripcija i prevođenje, au eukariota postoji i faza obrade.
Ekspresija gena se sastoji u sintezi molekule mRNA komplementarnoj (ili transkripciji - prepisivanju, čitanju bioloških informacija) na molekuli DNK i njegovoj daljoj upotrebi za sintezu proteina. Jedinica transkripcije u DNK je transkripton, koji je veći od strukturnih gena. Transkripton u eukariotskim ćelijama sastoji se od neinformativne (akceptorske) i informativne zone. Neinformativna zona počinje s genomom promotora (područje od 80 nukleotida), kojem enzim RNA polimeraza katalizira proces čitanja. U prokariotima, jedan tip RNA polimeraze, kod eukariota, tri.
Operativni geni koji vezuju regulatorne proteine (proteine koji uključuju i završavaju transkripciju) nalaze se iza promotorskog gena.
Informativna zona se sastoji od strukturnih gena koji se nalaze iza gena od strane operatora. Strukturni geni eukariota su odvojeni razmaknicama - DNK segmentima koji ne nose informacije.
Osim toga, struktura samih strukturnih gena uključuje informativne parcele - egzone i neinformativne - introne. Nemamo razmaknice, egzone i introne.
U svakoj fazi životnog ciklusa, samo 10% strukturnih gena se transkribuje u ćeliju, a preostali geni nisu aktivni, ali neki od njih mogu biti uključeni u druge faze životnog ciklusa.
Kao rezultat transkripcije, m-RNA (zrela RNK) se formira odmah u prokariotima i proces prevođenja počinje odmah.
Kod eukariota se transkribuje velika molekula mRNK koja sadrži sve neinformativne regione. Naziva se prekursor RNA ili pre-RNA. Stoga, transkripcija počinje da se obrađuje, zbog čega se uništavaju sve neinformativne sekcije: akceptorska zona, razmaknice i introni, a preostali egzoni se spajaju (spajanje).
U fazi obrade, nukleotidna sekvenca, nazvana kapica, se veže na početni segment formirane RNK, a sekvenca adeninskih ostataka, poli-A, se dodaje terminalnom segmentu.
U obradi, nukleotidi su modificirani u RNA, na primjer, njihova metilacija, hidrogenacija.
I tek nakon ovih transformacija formira se zrela m-RNK, koja počinje uvodnim redoslijedom, nazvanim vođa, a završava završnom sekvencom - prikolicom. Lider je uvodna nukleotidna sekvenca koja je komplementarna sekvenci u p-RNK molekulu male ribosomske podjedinice, koja osigurava vezivanje m-RNK za malu podjedinicu. Prikolica uključuje besmislen kodon i poli-A sekvencu.
Element koda m-RNK je nukleotidni trostruko nazvan kodon. Svaki kodon odgovara specifičnoj aminokiselini.
Primarna struktura RNK je, kao u DNK, linearni lanac polinukleotida, u kojima su mononukleotidi povezani 3 ', 5' -fosfoeter vezama.
Sekundarna struktura RNK je zakrivljeni lanac, a tercijarni je konac namotan na kalem, uloga zavojnice igra poseban transportni protein, doušnik.
Rezultirajuća mRNA ulazi u citoplazmu do mjesta sinteze polipeptida (proteina), tj. na ribosome.
Proces prevođenja se izvodi na ribosomima. Prevod je mehanizam kojim se sekvenca nukleotida u m-RNK molekulama prevodi u specifičnu aminokiselinsku sekvencu u polipeptidnom lancu.
Prevod se sastoji od tri faze:
1. Iniciranje - početak sinteze polipeptida.
2. Produženje - produljenje polipeptida.
3. Završetak - kraj sinteze polipeptida.
U početnoj fazi, manja podjedinica ribozoma prepoznaje startni kodon AUG m-RNK i pridaje mu se. AUG zauzima prvo mjesto. Nakon toga se spaja velika podjedinica ribozoma i počinje stvarna sinteza proteina. T-RNA sa amino kiselinom je pogodna za veliku podjedinicu.

U molekuli t-RNK, jedan deo dodaje aminokiselinu, a drugi, antikodon, parove sa kolonijom m-RNK, koja određuje ovu amino kiselinu.

U velikoj podjedinici postoje:
1 - aminoacil područje
2 - peptidil područje
U tRNA, čiji je antikodon komplementaran kodu AUG
m-RNA donosi aminokiselinu metianin i zaustavlja se na aminoacilnom mjestu. Faza inicijacije se kontroliše inicijacijskim faktorima. U prokariotima postoje 3, kod eukariota, 6-7.
Druga faza - izduženje počinje prenosom t-RNK sa metianinom na peptidil, a drugo t-RNK sa novom aminokiselinom zauzima svoje mesto. Peptidna veza se formira između dvije aminokiseline. U međuvremenu, ribozom se kreće duž m-RNK, pojavljuje se novi kodon na ribozomu, kome se odgovarajuća t-RNA uskoro pridružuje sa svojim antikodonom.
Sve ovo se ponavlja mnogo puta, dok ribosom nije
će doći do jednog od terminalnih kodona (UAA, UAG, UGA).
Ovi kodoni nemaju antikodon, nemaju aminokiseline. Dolazi do faze raskida. Mnogi su vezani za jedan molekul m-RNK.
formiraju se ribozomi i polisomi.
U procesu prevođenja, zajedno sa faktorima inicijacije i elongacije, važan je enzim koji veže t-RNK na aminoacil region, aminoacil t-RNAse.
Regulacija ekspresije gena kod pro- i eukariota
U prokariotima, on se izvodi na nivou promotora, operatora
i emitovanje.
Kod eukariota, regulacija se dešava u svim fazama ekspresije: transkripcija, obrada i translacija.
Da se zadržimo na regulaciji ekspresije gena u bakterijama.
Regulacija ekspresije gena u prokariotima

Šema regulacije transkripcije strukturnih gena prokariotskih ćelija prema tipu represije

Šema regulacije transkripcije strukturnih gena prokariotske ćelije tipom indukcije
Godine 1961. Jacob i Monod su otkrili da bakterije pod kontrolom regulacionog gena sintetišu represorski protein koji reguliše aktivnost drugih gena. Represorski protein vezuje se za operatora gena i blokira ga, što rezultira time da transkripcija (čitanje informacija) postaje nemoguća i da ćelija ne može sintetizirati odgovarajuće proteine. Ali ako bilo koji supstrat prodre u ćeliju, za cepanje koje je neophodno sintetizovati enzime, dešava se sledeće: supstrat se pridružuje represoru i lišava ga sposobnosti da blokira genskog operatora.
Enzimi koji sintetišu RNK su vezani za DNK i počinje transkripcija gena. Sinteza proteinskog enzima je prekinuta, ako je supstrat potpuno odcijepljen, represorski protein se oslobađa i ponovo blokira operatorske gene. Ovo je primjer regulacije indukcijskog tipa. Postoji i regulativa o vrsti represije: kada konačni proizvodi biohemijske reakcije, kombinujući se sa neaktivnim proteinskim represorom, formiraju kompleks koji blokira rad genskog operatora.
Antiterminacija - je da dolazi do ignorisanja terminalnih kodona, proces ekspresije se nastavlja.
Regulacija kod eukariota
1) Regulacija aktivnosti gena kod eukariota je mnogo komplikovanija nego kod bakterija. Kod eukariota se javlja ne samo na nivou ćelije. Postoje sistemi regulacije organizma u cjelini. Hormoni igraju veliku ulogu u regulaciji, ali reguliraju procese sinteze proteina samo u ciljnim stanicama. Hormoni se vezuju za receptorske proteine koji se nalaze u membranama takvih ćelija i uključuju sisteme za promenu strukture ćelijskih proteina. One, pak, mogu uticati na procese transkripcije i procese prevođenja. Svaki hormon kroz sistem posrednika aktivira svoju grupu gena. Dakle, adrenalin uključuje sintezu enzima koji razgrađuju glikogen mišića do glukoze, a insulin utiče na formiranje glikogena iz glukoze u jetri.
2) U fazi transkripcije, histonski proteini su uključeni u regulaciju aktivnosti gena kod eukariota. Neophodan uslov je dekondenzacija mjesta gdje se javlja transkripcija.
3) Regulacija na nivou prevoda ima za cilj očuvanje stabilnosti m-RNK, a efikasnost prevođenja se vrši pomoću faktora inicijacije, izduženja i završetka.
Predavanje 12.
Varijabilnost
Plan
1. Definicija i oblici varijabilnosti: modifikacija, kombinacija, mutacija.
2. Mutageni faktori.
3. Klasifikacija mutacija.
4. Održivost i metode reparacije genetskog materijala.
5. Zakon homologne serije nasljedne varijabilnosti N.A. Vavilova.
Definicija i oblici varijabilnosti
Genetika proučava dva glavna svojstva živih bića - nasleđe i varijabilnost.
Varijabilnost je svojstvo organizama za sticanje novih znakova i karakteristika individualnog razvoja pod uticajem faktora okruženja.
Varijabilnost je jedan od najvažnijih faktora evolucije, pružajući svu raznolikost žive prirode.
Postoje dvije vrste varijabilnosti.:
1. Fenotipski (ne-nasljedni, modifikacija);
2. Genotipski (nasljedni):
a) kombinacija;
b) mutacije.
Modifikacijska varijabilnost - oblik varijabilnosti, koji nije povezan sa promjenama u genotipu i uzrokovan utjecajem faktora okoline.
Varijabilnost modifikacije ima karakteristike:
Ne utiče na naslednu osnovu organizma i stoga se modifikacije ne nasleđuju, tj. Od roditelja do potomstva,
Promene su usmerene, javljaju se prirodno, mogu se predvideti
Imaju adaptivnu (adaptivnu) vrednost,
Imati masivnu (grupnu) i reverzibilnu prirodu,
Određeni ekološki faktor uzrokuje sličnu promjenu kod svih jedinki ove vrste.
Varijabilnost modifikacije ima granicu. Nazivaju se granice varijabilnosti svojstava zbog genotipa normalna reakcija. Može biti uska kada se znak neznatno promijeni (boja očiju), i široka, kada se znak znatno mijenja (visina, težina).
U medicini je često potrebno uspostaviti brzinu reakcije za procjenu maksimalnih i min kvantitativnih pokazatelja (nivo hormona, enzima, hemoglobina, itd.)
Kombinativna varijabilnost - ovo je nasljedna varijabilnost uzrokovana rekombinacijom postojećih gena i kromosoma, bez promjene strukture gena i kromosoma (nasljedni materijal). Ovaj tip varijabilnosti se manifestuje već u fazi formiranja zametnih ćelija.
Izvori kombinacione varijabilnosti su procesi koji se javljaju u mejozi i kao rezultat oplodnje:
1. Rekombinacija gena kod križanja u mezojima profaze 1.
2. Rekombinacija hromozoma tokom mejoze (nezavisna divergencija hromozoma i hromatida tokom mejoze)
3. Kombinacija hromozoma kao rezultat fuzije gameta tokom oplodnje (slučajna kombinacija gameta tokom oplodnje).
Kombinativna varijabilnost osigurava genotipsku raznolikost ljudi, objašnjava prisustvo znakova kod djece i unučadi od rođaka na roditeljskoj i majčinskoj liniji.
Mutacijska varijabilnost - sposobnost genetičkog (nasljednog) materijala da se promijeni i te promjene su naslijeđene u potomstvu.
Osnova mutacijske varijabilnosti su mutacije.
Mutacije su iznenadne promjene genetskog materijala pod utjecajem okoline i naslijeđene.
Učestalost mutacija zavisi od tipa organizma, od starosti, od faze ontogeneze, od faze gametogeneze, koja se može javiti u polnim i somatskim ćelijama, ima recesivan i dominantan karakter. Na primer, kod ljudi, do 6% gameta nosi mutantne gene.
Proces formiranja mutacija naziva se mutageneza.
Faktori koji uzrokuju mutacije nazivaju se mutageni.
Mutacije inicijalno djeluju na genetski materijal pojedinca, a fenotip se također mijenja kroz genotip.
Mutageni faktori
Faktori koji uzrokuju mutacije nazivaju se mutageni faktori (mutageni) i dijele se na:
1. Fizička;
2. Hemikalije;
3. Biološki.
Fizičkim mutagenim faktorima uključuju različite vrste zračenja, temperaturu, vlažnost, itd. Najsnažniji mutageni učinak je ionizirajuće zračenje - rendgenske zrake, α-, β-, γ-zrake. Oni imaju veliku moć prodiranja.
Kada djeluju na tijelo, uzrokuju:
a) ionizacija tkiva - formiranje slobodnih radikala (OH) ili (H) iz vode u tkivima. Ovi joni ulaze u hemijsku interakciju sa DNK, razlažu nukleinsku kiselinu i druge organske supstance;
b) ultraljubičasto zračenje karakteriše manje energije, prodire samo kroz površinske slojeve kože i ne uzrokuje ionizaciju tkiva, ali dovodi do formiranja dimera (kemijske veze između dvije pirimidinske baze jednog lanca, češće tt). Prisustvo dimera u DNK dovodi do grešaka u njegovoj replikaciji, narušava čitanje genetske informacije;
c) pucanje navoja vretena;
d) kršenje strukture gena i hromozoma, tj. formiranje genskih i hromozomskih mutacija.
Kemijski mutageni uključuju:
Prirodne organske i neorganske tvari (nitriti, nitrati, alkaloidi, hormoni, enzimi itd.);
Sintetičke supstance koje se ranije nisu susrele u prirodi (pesticidi, insekticidi, konzervansi hrane, ljekovite tvari).
Proizvodi industrijske prerade prirodnih spojeva - ugalj, nafta.
Mehanizmi njihovog djelovanja :
a) deaminacija - uklanjanje amino grupe iz molekula amino kiseline;
b) supresija sinteze nukleinske kiseline;
c) zamjena azotnih baza njihovim analozima.
Kemijski mutageni uzrokuju uglavnom mutacije gena i djeluju tokom perioda replikacije DNK.
Biološke mutagene uključuju:
Virusi (gripa, rubeola, ospice)
Mehanizmi njihovog djelovanja:
a) virusi ugrađuju svoju DNK u DNK ćelije domaćina.
Biološke mutagene uzrokuju mutacije gena i hromozoma.
Klasifikacija mutacija
Razlikuju se sljedeće glavne vrste mutacija:
1. Kao pojava podeljeni su na spontane i indukovane.
Spontano- javljaju se pod uticajem prirodnih mutagenih faktora okoline bez ljudske intervencije. Oni nastaju u uslovima prirodne radioaktivne pozadine Zemlje u obliku kosmičkog zračenja, radioaktivnih elemenata na površini Zemlje.
Induced mutacije su veštački indukovane određenim mutagenim faktorima.
2. Od mutiranih ćelija mutacije se dijele na generativne i somatske.
Generative- javljaju se u zametnim ćelijama, nasleđuju se tokom seksualne reprodukcije.
Somatic- pojavljuju se u somatskim ćelijama i prenose se samo na one ćelije koje nastaju iz ove somatske ćelije. Oni se ne nasleđuju.
3. Uticajem na telo:
Negativne mutacije su smrtonosne (nespojive sa životom); poluletalni (smanjenje vitalnosti organizma); neutralna (ne utiče na procese vitalne aktivnosti); pozitivno (povećanje održivosti). Pozitivne mutacije su rijetke, ali su važne za progresivnu evoluciju.
4. Prema genetskim promenama Materijalne mutacije su podijeljene na genomske, kromosomske i genske.
Genomske mutacije - To su mutacije uzrokovane promjenom broja kromosoma. Mogu se pojaviti ekstra homologni hromozomi. U setu hromozoma na mestu dva homologna hromozoma postoje tri - to je trisomija. U slučaju monosomije, postoji gubitak jednog hromozoma iz para. Kada se poliploidija javi višestruko više od haploidnog povećanja broja hromozoma. Druga varijanta genomske mutacije je haploidija, u kojoj ostaje samo jedan hromozom iz svakog para.
Chromosomalmutacije su povezane sa kršenjem strukture hromozoma. Takve mutacije uključuju gubitak kromosomskih regija (delecija), dodavanje regija (duplikacija) i rotaciju regije kromosoma za 180 ° (inverzija).
Genesmutacije u kojima se promene dešavaju na nivou pojedinačnih gena, tj. sekcije DNK molekula. To može biti gubitak nukleotida, zamjena jedne baze drugom, preraspodjela nukleotida ili dodavanje novih.
Otpornost i reparacija genetskog materijala
Otpornost na promjene u genetskom materijalu osigurava:
1. Delfidni set hromozoma.
2. DNK dvostrukog heliksa.
3. Degeneracija (redundancija) genetskog koda
4. Ponavljanje nekih gena.
5. Popravljanje poremećaja strukture DNA
Postojanje mehanizama reparacije je neophodan uslov za postojanje bioloških bića.
Reparacija genetskog materijala – To je proces koji osigurava obnavljanje oštećene strukture DNK molekula.
Mnogo nasumičnih promena se dešava svakog dana u DNK ćelije.
Većina se efikasno koriguje (popravlja) uz pomoć posebnih enzimskih sistema.
Prvi popravak DNK uspostavljen je 1948. godine. Godine 1962. opisan je jedan od načina reparacije. - popravljanje ili fotoreaktivacija.
Utvrđeno je da se ultraljubičastim zračenjem virusa faga, bakterija i protozoa uočava naglo smanjenje njihove vitalne aktivnosti, čak i smrti.
Ako na njih utičete vidljivom svetlošću, njihova stopa preživljavanja se značajno povećava.
Pokazalo se da se pod dejstvom ultraljubičastog zračenja u molekuli DNK formiraju dimeri (hemijske veze između dve baze istog lanca, češće T-T), formiranje dimera sprečava čitanje informacija.
Vidljiva svetlost aktivira enzime koji razbijaju dimere.
Druga metoda reparacije - mračna reparacija, proučavana je 50-ih godina dvadesetog veka.
Dark reparationnastavlja se u četiri faze uz učešće četiri grupe enzima. Enzimi su nastali tokom evolucije i imaju za cilj održavanje stabilnosti genetske informacije ćelije.
1. Enzim endonukleaza pronalazi oštećenu površinu i pored nje lomi DNK lanac.
2. Enzim ektonukleaza "izrezuje" (uklanja) oštećeno područje.
3. Komplementarnost DNA polimeraze sintetizira fragment DNA na mjestu uništenog.
4. Ligaza "šiva" sintetizirani fragment s glavnim lancem DNA.
Dokazana je mogućnost obnove DNK u slučaju oštećenja oba niti. Istovremeno se mogu dobiti informacije sa i-RNA (enzimom revertaze).
Zakon homologne serije nasljedne varijabilnosti N. Vavilova
Poznato je da se mutacija odvija u različitim pravcima. Međutim, ova raznolikost je podložna određenoj pravilnosti, koju je 1920. otkrio N.Vavilov. On je formulisao zakon homologne serije nasledne varijabilnosti: "Vrste i rodovi, genetski bliski, karakterizirani su sličnim nizom nasljedne varijabilnosti s takvom točnošću da poznavanje broja oblika unutar jedne vrste može predvidjeti postojanje paralelnih oblika kod drugih vrsta i rodova."
Može se reći da srodne vrste koje imaju zajedničko porijeklo imaju slične mutacije.
Iste bolesti se javljaju kod životinja i ljudi. Na primer, albinizam je primećen u svim klasama životinja i ljudi. Brachydactyly (kratkoća) je uočena kod štakora, ovaca, pasa, ljudi. Mišićna distrofija - kod miševa, pacova, konja, ljudi.
Zakon homologne serije omogućava da se predvidi mogućnost pojave mutacija koje se mogu koristiti u selekciji za stvaranje novih oblika koji su dragoceni za ekonomiju.
Metode proučavanja ljudske genetike
Plan
1. Genealoška metoda.
2. Dvostruka metoda.
3. Metoda dermatoglifika.
4. Citogenetička metoda.
5. Metoda hibridizacije somatskih ćelija.
6. Ontogenetska metoda.
7. Populacijsko-statistička metoda.
8. Metoda modeliranja.
9. Imunološka metoda.
10.Biohemijska metoda.
Genealoška metoda
Vrste nasljeđivanja i oblici ispoljavanja genetskih sklonosti u osobi su vrlo različiti i potrebne su posebne metode analize za njihovo razlikovanje, prije svega, genealoška metoda koju je predložio F. Galton.
Genealoška metoda ili proučavanje rodovnika predviđa praćenje osobine u porodici ili rodu, ukazujući na vrstu srodstva između članova pedigrea. U medicinskoj genetici, ovaj metod se obično naziva kliničkim i genealoškim, jer se radi o posmatranju patoloških znakova koristeći tehnike kliničkog pregleda. Genealoška metoda se odnosi na univerzalne metode u ljudskoj genetici. Široko se koristi u rješavanju teorijskih i praktičnih problema:
1) da se utvrdi nasledna priroda te osobine,
2) u određivanju vrste nasleđivanja i penetracije genotipa,
3) detekcija povezivanja gena i mapiranje hromozoma,
4) prilikom proučavanja intenziteta procesa mutacije,
5) kod dešifriranja mehanizama interakcije gena,
6) sa medicinskim i genetskim savetovanjem.
Suština genealoške metode svodi se na razjašnjavanje srodstva i pronalaženje osobine među bliskim i daleko direktnim i indirektnim rođacima. Tehnički, sastoji se od dvije faze: kompilacije pedigree i genealoške analize.
Kompilacija pedigrea
Prikupljanje informacija o porodici započinje probandom, koji je prva osoba u vidnom polju istraživača.
Djeca jednog roditeljskog para (braća i sestre) se nazivaju braća i sestre. Porodica u užem smislu, ili nuklearna porodica, zove se roditeljski par i njihova djeca. Širi krug krvnih srodnika bolje je označen terminom "rod". Što je više generacija uključeno u pedigre, to je to opsežnije. To podrazumijeva netočnost primljenih informacija i, shodno tome, netočnost pedigrea u cjelini. Često ljudi ne znaju ni broj svojih rođaka i sestara, a kamoli neke znakove njih i njihove djece.
Radi jasnoće, priprema grafičke slike pedigrea. Za to se obično koriste standardni znakovi. Ako su razmatrani znakovi u pedigreu mnogo, onda možete pribjeći razlikama slova ili moždanog udara unutar znakova. Pedigree shema je nužno popraćena opisom notacije ispod slike - legenda, koja isključuje mogućnost pogrešnog tumačenja.
Genealoška analiza
Cilj genealoške analize je utvrđivanje genetskih obrazaca.
Faza 1 – utvrđivanje nasledne prirode ove osobine. Ako se u pedigreu jedan i isti znak pojavljuju nekoliko puta, onda se može misliti na njegovu naslednu prirodu. Međutim, prvo moramo isključiti mogućnost egzogene akumulacije slučajeva u porodici ili rodu. Na primer, ako je isti patogeni faktor uticao na ženu tokom svih trudnoća, onda bi mogla imati nekoliko dece sa istim anomalijama. Ili je neki faktor djelovao na mnoge članove porodice, potrebno je uporediti akciju sličnog vanjski faktori. Uz pomoć genealoške metode opisane su sve nasljedne bolesti.
Faza 2 – određivanje tipa nasleđivanja i penetracije gena. U tu svrhu koriste se principi genetske analize i statističke metode za obradu podataka iz pedigrea.
Faza 3 – određivanje grupa za povezivanje i mapiranje hromozoma, do nedavno zasnovano samo na genealoškoj metodi. Otkrijte povezane funkcije i crossover proces. Ovo je olakšano razvijenim matematičkim metodama.
Faza 4 – proučavanje procesa mutacije. Primjenjuje se u tri smjera: u proučavanju mehanizama mutacije, intenzitetu mutacijskog procesa i faktora koji uzrokuju mutacije. Genealoška metoda se posebno široko koristi u proučavanju spontanih mutacija, kada je potrebno razlikovati „sporadične“ slučajeve koji su nastali iz „porodice“.
5 faza – analizu interakcije gena u kliničkoj genetici izvršio je S.N.Davidenkov (1934, 1947) na analizi polimorfizma bolesti nervnog sistema.
6 faza – u genetičkom savjetovanju za pripremu prognoze nemoguće je bez genealoške metode. Oni otkrivaju homo- ili heterozigotnost roditelja i razmatraju vjerovatnoću da imaju djecu sa određenim znakovima.
Metod dvostruke studije
Proučavanje blizanaca - jedna od glavnih metoda ljudske genetike. Postoje identični blizanci koji nastaju iz jednog jajeta oplođenog jednom jedinom spermatozoidom. Oni nastaju usled razdvajanja zigota na dva genetski identična i uvek istospolnog embriona.
Bratski blizanci nastaju iz različitih jaja oplođenih različitim spermatozoidima. Genetski se razlikuju kao braća i sestre istih roditelja.
Upotrebom dvostruke metode može se proučavati:
1) Uloga nasleđa i životne sredine u formiranju fizioloških i patoloških osobina organizma. Konkretno, proučavanje nasljedne transmisije određenih bolesti od strane ljudi. Proučavanje ekspresivnosti i penetracije gena koji uzrokuju nasljedne bolesti.
2) Specifični faktori koji pojačavaju ili slabe uticaj spoljašnjeg okruženja.
3) Korelacija karakteristika i funkcija.
Posebno je važna uloga dvostruke metode u proučavanju problema "genotipa i okoline".
Obično se uspoređuju tri grupe blizanaca: DB u istim uvjetima, OB u istim uvjetima, OB u različitim uvjetima.
U proučavanju blizanaca određuje se učestalost, stepen podudarnosti (podudarnosti) određenih znakova.
Proučavajući ulogu nasleđa u poreklu određene osobine, izračun se vrši prema formuli K. Holzinger.
Koeficijent heritabilnosti - N
H =% sličnosti OB -% sličnosti RB
100% sličnost RB
Kada je H = 1, sva varijabilnost u populaciji je posljedica nasljednosti.
Kada je H = 0, sva varijabilnost je uzrokovana faktorima okoline. Uticaj medija C izražava se formulom: C = 100% - H, gdje je H koeficijent heritabilnosti. Na primer, podudarnost monozigotnih (identičnih) blizanaca je 3%. Zatim H =
67 – 3 = 64 = 0,7 ili 70%. C = 100 - 70 = 30%
Dakle, ova osobina je 70% zbog nasljednosti, a 30% zbog utjecaja okolišnih faktora.
Još jedan primjer. Krvne grupe u ABO sistemu kod OB = 100%, tj. potpuno zavisna od naslednosti.
Učestalost podudarnosti krvnih grupa i određenih bolesti kod blizanaca (u%)
|
Znakovi ili bolesti | |||
|
aBO krvne grupe | |||
|
šizofrenija | |||
|
epilepsija | |||
|
Kongenitalna stenoza pilorusa |
Dermatoglifska metoda
Uhto je nauka koja proučava nasljednu uslovljenost obrazaca koji formiraju linije kože na vrhovima prstiju, dlanovima i tabanima osobe.
Pokazalo se da svaka nacija, svaka rasa, svaka osoba ima svoje obrasce, i da su strogo individualne na dlanovima. F. Galton je prvo skrenuo pažnju na to, koji je predložio britanskoj kriminalističkoj policiji da identifikuje kriminalce pomoću otisaka prstiju.
Dermatoglifske studije su važne u forenzičkoj nauci, u određivanju zigoznosti blizanaca, u dijagnostici niza nasljednih bolesti, kao iu nekim slučajevima kontroverznog očinstva.
Reljef palme je veoma složen. Razlikuje niz polja, jastučića i dlanovnika. Jastuci na dlanu 11, podijeljeni su u 3 grupe:
1) pet krajnjih (eplical) jastučića na krajnjim falangama prstiju.
2) četiri interdigitalna jastučića, smještena protiv interdigitalnih prostora.
3) dva palmarna proksimalna jastučića tenora i hipotenara. U bazi thumb - Tenor, na suprotnoj ivici dlana-hipotenara.
Na najuzvišenijim delovima jastučića vidljive su kožne kapice. To su linearno zgušnjavanje epidermisa, koje su modifikovane skale kože. Školjka ide u potocima, i na dlanovima i na vrhovima prstiju. Točke susreta ovih potoka formiraju triradii ili delte.
Šuplji uzorci se obično proučavaju pod povećalom. Štampa uzoraka, koristeći štampanje tinte, učiniti na čistom bijelom, bolje obložene, papir ili celofan. I na vrhovima prstiju i na palmarskim uzvišenjima, mogu se uočiti različiti papilarni obrasci u obliku uvojki, petlji i lukova otvorenih na ulcerativnim ili radijalnim stranama. Na teneru i hipotenaru češće postoje lukovi. Na srednjim i glavnim falangama prstiju, linije poput češlja prolaze kroz prste, formirajući različite obrasce - ravne, srpaste, talasaste, lučne i njihove kombinacije. U proseku, 15-20 morskih kapica su na jednom prstu.
Figure palm.

1 - poprečni proksimalni žlijeb, linija pritiska 4 prsta
2 - poprečna srednja brazda, linija presovanja od 3 prsta
3 - poprečni distalni žlijeb, linija za pritisak 2 prsta
4 - utor za palac
5 - uzdužni srednji žlijeb od zapešća do baze 3. prsta
6 - uzdužni srednji žlijeb od ručnog zgloba do podnožja 4. prsta
7 - uzdužni žljeb ulnara, od ručnog zgloba do baze petog prsta

1 - Patau sindrom
2 - Downov sindrom
3 - Shereshevsky-Turner-ov sindrom
4 - norma
5 - Klinefelterov sindrom
U istraživanju reljefa kože dlana istražite:
1) Hod glavnih linija dlana A, B, C, D 1,2,3,4,5,6,7.
2) Dlanovi na teneru i hipotenaru.
3) Uzorci prstiju (oblik uzoraka, broj grebena)
4) aksijalni triradii.
Slične studije se izvode na tabanima. Pravac glavnog dlana D je isti za roditelje i njihovu decu.
Ispitivanje pacijenata sa hromozomskim bolestima (Downova bolest, Klinefelter sindrom) pokazalo je da oni menjaju ne samo uzorak prstiju i dlanova, već i prirodu glavnih žljebova na koži dlanova.
Dermatoglifske abnormalnosti kod pacijenata sa takvim razvojnim defektima kao što su kongenitalni defekti srca i velike krvne sudove, nekonjugacija mekog i tvrdog nepca, gornje usne, itd.
Utvrđene su promjene u karakteru oblika prstiju i dlana kod gube, šizofrenije, dijabetesa, raka, reumatizma, dječije paralize i drugih bolesti.
Citogenetička metoda
Ova metoda omogućava da se pomoću mikroskopa istraži struktura ćelije - hromozoma. Metodom mikroskopije proučavan je kariotip ljudskog tijela (skup kromosomskih ćelija tijela). Utvrđeno je da su mnoge bolesti i razvojni defekti povezani sa kršenjem broja hromozoma i njihove strukture. Ova metoda omogućava i proučavanje efekta mutagena na sastav i strukturu hromozoma. Citogenetska metoda je povezana sa privremenim tkivnim kulturama (obično leukociti) i proizvodnjom metafaznih jezgara sa skraćenim, zadebljanim hromozomima, čija se podjela prekida na stadiju metafazne ploče pomoću kolhicina. Ako se hromozomi proučavaju u kariotipu, onda ovaj metod omogućava istraživanje spolnog kromatina u somatskim ćelijama.
Hibridizacija somatskih ćelija
Hibridne ćelije imaju određena svojstva koja vam omogućuju da odredite lokalizaciju genske ili genske veze. Gubitak ljudskih hromozoma iz nekih tipova hibridnih ćelija omogućava dobijanje klonova u odsustvu specifičnog hromozoma. Najčešće korišćeni hibridi ljudskih somatskih ćelija su miš.
Praćenje prisustva biohemijskog genetskog markera u hibridnim klonovima kao eliminaciji humanih hromozoma može dovesti do otkrivanja lokalizacije gena, ako ta osobina nestane iz ćelija čim promene određene hromozome. Citogenetička analiza velikog broja klonova i poređenje rezultata sa prisustvom velikog broja genetskih markera omogućava nam da uočimo povezane gene i njihovu lokalizaciju. Osim toga, informacije se koriste kada se koriste klonovi od osoba sa invaliditetom sa translokacijama i drugim kromosomskim abnormalnostima.
Ovaj metod je korišćen za lociranje gena fosfoglicerat kinaze u dugom kraku X hromozoma, tj. stavite hibridne ćelije omogućava vam da instalirate:
1) lokalizacija gena
2) povezivanje gena
3) mapiranje hromozoma
Više od 160 lokusa identificirano je metodom hibridnih somatskih stanica.
Ontogenetska metoda
Omogućava vam da proučite obrasce manifestacije bilo kojeg znaka ili bolesti u procesu individualnog razvoja. Postoji nekoliko perioda ljudskog razvoja. Antenatalni (razvoj prije rođenja) i postnatalni. Većina ljudskih osobina formira se u fazi morfogeneze antenatalnog perioda. U fazi morfogeneze postnatalnog perioda, završava se formiranje moždane kore i nekih drugih tkiva i organa, formira se imunološki sistem tela, koji dostiže svoj najveći razvoj 5-7 godina nakon rođenja djeteta. U postmorfogenetskom periodu razvijaju se sekundarne spolne karakteristike.
U morfogenetskom periodu dolazi do promjene aktivnosti gena u dvije vrste:
1) uključivanje i isključivanje gena
2) jačanje i slabljenje djelovanja gena
U postmorfogenetskom periodu razvoja, prvi tip promjene aktivnosti gena je gotovo odsutan, postoji samo malo uključivanje pojedinih gena - na primjer, geni koji određuju sekundarne spolne karakteristike, razvoj određenih nasljednih bolesti. Isključivanje gena u ovom periodu je značajnije. Aktivnost mnogih gena povezanih s proizvodnjom melanina je potisnuta (kao rezultat toga dolazi do sijanja), kao i gena povezanih s proizvodnjom γ-globulina (osjetljivost na bolesti se povećava). Mnogi geni su potisnuti u ćelijama nervnog sistema, mišićnih ćelija itd.
Represija gena se dešava na nivou transkripcije, prevođenja, post-translacije. Međutim, glavni tip promjene aktivnosti gena u ovoj fazi je jačanje i slabljenje djelovanja gena. Dominacija gena može se promijeniti, što uzrokuje promjenu vanjskih znakova, posebno tijekom puberteta. Menja odnos spolnih hormona i, shodno tome, znakova seksa. Represivni geni sa godinama mogu imati veliki uticaj na razvoj određene osobine. Na primjer, fenilketonurijski gen u heterozigotnom stanju mijenja ljudsku psihu.
Metoda populaciono-statističkih istraživanja
To je metoda matematičkog proračuna različitih gena i odgovarajućih osobina u određenim populacijama. Teorijska osnova ove metode je Hardy-Weinbergov zakon.
Ova metoda je utvrdila da se svi geni ljudske populacije mogu podijeliti u 2 kategorije prema učestalosti pojavljivanja:
1) da ima univerzalnu distribuciju kojoj pripada većina gena. Na primer, gen za slepilo za boje dostupan je kod 7% muškaraca i više od 13% žena. Gen za amavrotičku idiotizam, pronađen u populaciji Evrope sa učestalošću od 4 na 10.000 ljudi.
2) geni koji se nalaze uglavnom u određenim područjima. Na primer, gen anemije srpastih ćelija je uobičajen u zemljama gde je malarija raširena. Gene kongenitalna dislokacija kuka, koja ima visoku koncentraciju kod stanovnika severoistoka naše zemlje.
Metoda simulacije
Zakon homologne serije N.Vavilova (genetski bliske vrste i rodovi imaju sličan niz nasljedne varijabilnosti) omogućava ekstrapolaciju eksperimentalnih podataka na ljude sa određenim ograničenjima.
Biološki model nasljedne bolesti kod životinje je često pogodniji za istraživanje nego bolesna osoba. Pokazalo se da životinje imaju oko 1300 nasljednih bolesti, kao i kod ljudi. Na primer, kod miševa - 100, kod krokodila - 50, kod pacova - 30. Na modelu hemofilije A i B kod pasa, pokazalo se da je to posledica recesivnog gena lociranog na X hromozomu.
Modeliranje mišićne distrofije kod miševa, hrčaka i pilića omogućilo je razumijevanje patogenetske prirode ove bolesti. Utvrđeno je da kod ove bolesti nije zahvaćen nervni sistem, već mišićna vlakna sama.
Početni mehanizmi galaktosemije su razjašnjeni na modelu Escherichia coli. I kod ljudi i kod bakterija, nemogućnost asimilacije galaktoze je uzrokovana istim nasljednim defektom - odsustvom aktivnog enzima - galaktoza-1-fosfatil uridil transferaze.
Metoda imunološkog istraživanja
Ova metoda se zasniva na proučavanju antigenog sastava ćelija i telesnih tečnosti ljudskog organizma - krvi, pljuvačke, želučanog soka itd. Najčešći antigeni su krvne stanice: crvena krvna zrnca, bela krvna zrnca, trombociti i krvni proteini. Različiti tipovi eritrocitnih antigena formiraju sisteme krvnih grupa.
Početkom dvadesetog veka K. Landsteiner i Yansky pokazali su da, u zavisnosti od prirode reakcija između eritrocita i krvne plazme, svi ljudi mogu biti podeljeni u 4 grupe. Kasnije je dokazano da se te reakcije javljaju između proteinskih supstanci eritrocita, koje se nazivaju aglutino-geni, i serumskih proteina, koji su nazvani aglutinini.
Grupe krvi određuju antigeni koji sadrže lipidne i proteinske frakcije, a nalaze se na površini eritrocita. Proteinski dio antigena kontrolira gen koji djeluje u ranim fazama razvoja eritrocita. Antigeni su specifični za svaku krvnu grupu.
Ukupno je poznato 14 sistema krvnih grupa eritrocita, koji uključuju više od 100 različitih antigena. U sistemu ABO krvnih grupa, na površini eritrocita pod kontrolom genskih alela formiraju se dva antigena. I a , I u .
Bernstein je 1925. godine pokazao da postoji treći alel I o koja ne kontrolira sintezu antigena. Dakle, postoje tri alela u sistemu ABO krvnih grupa, ali svaka osoba ima samo dva. Ako naslikamo moguće muške i ženske gamete u Punnettovoj rešetki, onda možemo pratiti moguće kombinacije krvnih grupa u potomcima.
ABO krvne grupe u potomcima u zavisnosti od krvnih grupa roditelja
Imunološke metode koje se koriste za procjenu pacijenata i njihovih rođaka za koje se sumnja immunodefetsitnye stanju (agammaglobulinemia, dysgammaglobulinemia, ataksija-telangiectatica, i drugi), kako se sumnja, antigen nespojivosti majke i fetusa, uz orgulje i tkiva transplantacija, kada se uspostavljaju prave veze, u slučajevima medicinski genetsko savjetovanje, kada je potrebno proučavati genetske markere u dijagnosticiranju povezanosti gena ili određivanju nasljedne osjetljivosti na bolesti, Prilikom uspostavljanja zygosti blizanaca.
Određivanje članstva u krvnim grupama je od praktične važnosti u različitim genetskim studijama:
1) prilikom uspostavljanja zigotnih blizanaca
2) u uspostavljanju povezanosti gena.
3) u forenzičkom pregledu u slučajevima kontroverznog očinstva ili materinstva. Poznato je da dijete nije moglo imati antigene koji nisu u roditeljima.
Sistem M krvne grupe otkrio je K. Landsteiner i I. Levin 1927. godine (u ovoj grupi se ne proizvode antitela na odgovarajuće antigene). Sistem ima dva alela M, N.
Geni koji određuju faktor M i N su kodominantni, tj. ako se sastanu zajedno, oboje se pojavljuju. Dakle, postoje homozigotni genotipovi MM i NN, te heterozigotni MN. U evropskim populacijama, genotipovi MM nalaze se u približno 36%, NN u 16%, a MN u 48%.
I geni, respektivno:
M = 36 + 48 = 60%
Rhesus factor
Kao što pokazuju istraživači, 85% Evropljana ima antigen eritrocita, koji je uobičajen kod antigena majmuna vrsta rezus majmuna. 15% ljudi nema Rh antigene na površini eritrocita.
Sistem grupa antigena Rh je veoma složen. Pretpostavlja se da se Rh antigeni kontrolišu sa tri blisko povezana lokusa C, D i E u dva hromozoma i dominantno nasleđuju. Stoga su za svaki lokus moguća tri genotipa: homozigotni Rh-pozitivni, heterozigotni Rh-pozitivni i homozigotni Rh-negativni.
Najimunogeniji je antigen D . Antigeni C i E su manje aktivni.
Godine 1962. utvrđeno je prisustvo eritrocitnog izoantigena X d koji se prenosi kroz hromozom spola X. \\ t Prema ovom antigenu, svi ljudi se mogu podijeliti na X-pozitivne i X-negativne. Među X d-pozitivnim ženama pronađeno je 88%, a među muškarcima - 66%. Ako su oba roditelja XD-negativna, onda će sva njihova djeca (djevojčice i dječaci) biti X-negativna. Ako je otac Xd-pozitivan, a majka je Xd-negativna, onda će njihove kćerke biti X-pozitivne, a sinovi X-negativni. Ako je majka X d-pozitivna, a otac X d-negativan, onda će njihovi sinovi biti X-pozitivni. tip nasljeđivanja križnog tipa. Kćerke mogu biti Xd-pozitivne ili Xd-negativne, zavisno od homozigotnosti majke. Gene X d - grupa je lokalizovana u kratkom kraku X hromozoma. XD sistem se koristi za proučavanje aneuploidije (abnormalan broj X hromozoma kod deteta sa trisomijom X, Klinefelterovim sindromom, Shereshevsky-Turner sindromom, itd.). Pretpostavlja se da Xd-inkompatibilnost majke i fetusa (majka Xd-negativna i fetus Xd-pozitivna) dovodi do smanjenja učestalosti rađanja djevojčica.
Biohemijska metoda
Omogućava, s jedne strane, da proučava količinu DNK u ljudskim ćelijama u zdravlju i bolesti, s druge strane, da odredi nasljedne metaboličke defekte uz pomoć:
1) određivanje abnormalnog proteina (strukturnih proteina ili enzima) koji nastaju kao rezultat biohemijskih reakcija;
2) definicija srednjih metaboličkih proizvoda koji se javljaju kao rezultat genetskog bloka reakcije direktne razmene.
Na primjer, u fenilketonuriji, aminokiselina fenilalanin se ne pretvara u tirozin. Povećava se koncentracija u krvi i smanjenje koncentracije tirozina. U ovom slučaju, fenilalanin se pretvara u fenilpiruvičnu kiselinu i njene derivate - fenil-mleko, fenilacetat i fenilacetilglutamin.
Ova jedinjenja se detektuju u urinu pacijenta koristeći FeCl 3 ili 2,4-dinitrofenilhidrazin feri hlorid.
Jedan od fenomena koji modifikuje klasičnu podelu prema fenotipu 9: 3: 3: 1 u hibridnoj hibridizaciji je nepotpuna dominacija.
Na primjeru monohibridnog prelaza u noćnoj ljepoti (Mirabilis jalapa) i drugih objekata, pokazano je da se s nepotpunom dominacijom jednog od alela gena pojavljuje hibrid prve generacije u ispoljavanju alternativnih osobina. U potomstvu takvog hibrida u F 2, razdvajanje fenotipa se odvija u odnosu 1: 2: 1, što odgovara razdvajanju genotipa. Činjenica da se u ovom slučaju heterozigotni potomci razlikuju izgled iz homozigota, omogućava nam da zaključimo da je dominacija rezultat interakcije para alela koji određuju razvoj ove osobine. Drugim riječima, dominacija odražava prevlast jednog alela gena nad drugim (na primjer, A preko a, B iznad b, itd.). Očigledno, takva interakcija članova alelnog para ne dovodi uvek do potpune dominacije, odnosno potpune dominacije.
Strogo govoreći, vjerovatno je da geni sa potpunom dominacijom uopće ne postoje, jer gen, kao što ćemo vidjeti kasnije, ne određuje niti jednu određenu osobinu ili svojstvo, već utječe na brojne osobine. Stoga, govoreći o dominaciji ili recesiji bilo kog gena, uslovno pratimo samo jednu stranu njegove akcije - razvoj jedne osobine.
U dihibridnom križanju sa nepotpunom dominacijom dva para alela, razdvajanje fenotipa u F 2 odgovara formuli za razdvajanje (1: 2: 1) 2, ili 1: 2: 2: 4: 1: 2: 1: 2: 1. Tako ćemo u F 2 9 imati fenotipske klase, što odgovara broju genotipskih klasa - 3 2. Navedimo dva primjera hibridnog hibridnog križanja s nepotpunom dominacijom dva para alela u životinji (piletina) i biljci (jagoda).
Kod pilića je poznat gen koji uzrokuje snažno kovrčavo perje. Alel gene ovog gena nepotpuno dominira njenim normalnim alelom a, koji određuje normalnu strukturu olovke. Tu je još jedan alelni par, koji se nalazi na drugom paru homolognih hromozoma, što uzrokuje takozvanu boju pljuskanog perja - crne tačke na beloj pozadini. Ovu mrlju kontrolira nepotpuno dominantan alel Ḃ. Recesivni alel b - određuje crnu boju. Prilikom prelaska kokoške sa snažnom kovrčavošću i crnim perjem Ãbbb sa pijetlom koji je poprskao boju i normalno aa perje, prva generacija hibrida ÃḂb-a se pokazala kao posredna u izražajnosti ovih znakova: oni pokazuju slabu kovrčavost i plavu boju. U drugoj generaciji takve hibridizacije hibridizacije, fenotipska podjela se ne vidi u omjeru 9: 3: 3: 1, već u omjeru 1: 2: 2: 4: 1: 2: 1: 2: 1.
Drugi primjer može se navesti iz eksperimenata TS Fadeeva, provedenih analizom nasljeđivanja karakteristika boje jagodičastog voća i oblika čašice u jagodama (Fragaria vesca). Crvena boja jagodastog voća određena je homozigotnim stanjem alela, bijelim - alelom a. Drugi alelni par u homozigotnom stanju određuje normalnu strukturu čašice ḂḂ, a recesivno homozigotno stanje bb gena dovodi do formiranja čaške u obliku lista.
Prva generacija hibridnih biljaka pervogoaḂb ima ružičaste bobice i srednju strukturu čašice. Od samoprašivanja takvih oblika u drugoj generaciji, razvijaju se biljke koje čine 9 različitih fenotipskih klasa, što odgovara 9 klasa genotipova. Koincidencija teorijski očekivane podele prema fenotipu sa onom koja je uočena u eksperimentu potvrđena je statistički: χ 2 = 11,37, 0,20\u003e P\u003e 0,10.
Češće se takvi slučajevi susreću kada se nepotpuna dominacija javlja samo u jednom od dva para alela u hibridnoj hibridizaciji. Klasičan primjer u tom smislu je podjela u F 2 prema boji i obliku cvijeta u snapdragonu (Antirrhinum majus). Crvena boja cvijeta u lavljem ždrijelu određena je dominantnim alelom, sa nepotpunom dominacijom nad recesivnim alelom, koji određuje bijelu boju. Drugi par znakova odnosi se na oblik cvijeta: normalan - dominantan i pyloric - recesivan.

U ovom hibridnom križanju, razdvajanje u F 2 daje 6 fenotipskih klasa: 3: 6: 1: 2: 3: 1, sa crvenim cvetovima samo u slučaju homozigotnog stanja dominantnog alela gena za bojenje, ružičastog cveta njegovog heterozigotnog stanja i homozigotnog stanja recesivni alel ovog gena. U obliku cvijeta neće biti intermedijarnih oblika, jer se heterozigotni i homozigotni dominantni oblici ovdje ne razlikuju.
Dakle, nepotpuna dominacija nad jednim ili oba para alela gena u dihibridnom križanju mijenja podjelu u fenotip. Uz punu dominaciju u F 2, pojavit će se 4 fenotipske klase, s nekompletnom dominacijom u jednom od alelnih parova gena - 6 i sa nepotpunom dominacijom u oba alelna para - 9. U posljednjem slučaju, cijepanje fenotipom potpuno se podudara s cijepanjem po genotipu.
Neophodno je ponoviti ranije izjavu da nepotpuna dominacija, iako komplicira sliku Mendelovog razdvajanja u dihibridnom prelazu, ni na koji način ne proturječi Mendelovim zakonima, već ih potvrđuje. Fenomen nepotpune dominacije otkriva novi sadržaj zakona nasleđivanja. Ako se slažemo s činjenicom da je ta osobina uzrokovana genomom i da je priroda razdvajanja u F 2 određena ponašanjem kromosoma u mejozi, onda obilježja cijepanja s nepotpunom dominacijom na nov način otkrivaju učinak gena i prirodu njegove manifestacije. Priroda manifestacije gena takođe može da promeni rascep fenotipa.
Dakle, fenomen nezavisnog razdvajanja može se izvršiti pod sljedećim uvjetima:
- pronalaženje gena koji se broje u nehomolognim hromozomima; njihov broj ne može premašiti haploidni broj hromozoma kod ove vrste;
- jednako vjerovatna formacija gameta svih vrsta na bazi slučajne divergencije kromosoma u mejozi;
- izjednačeno sazrevanje gameta svih vrsta;
- izjednačen susret gameta tokom oplodnje;
- jednako vjerovatno preživljavanje zigota i odraslih organizama;
- relativna stabilnost ispitivanih osobina. Ali svi ovi uslovi nastaju pod uticajem raznih, konstantno i privremeno delujućih faktora. Zbog toga, oni mogu da se promene, menjajući ispoljavanje obrazaca nasleđivanja.
Ako pronađete grešku, označite dio teksta i kliknite Ctrl + Enter.