Šta je recesivni gen? Recesivni geni
Heredity - \\ tsposobnost organizama da ponavljaju slične znakove među generacijama. Roditelji i potomci imaju sličan izgled, fizičku strukturu, metabolizam. Kao rezultat, svaka vrsta se reprodukuje iz generacije u generaciju.
Nasljednost je rezultat kombinacija gena.
Genes - to su biohemijski funkcionalni elementi hromozoma koji određuju potencijalne polne i druge znakove embriona.
Hromozomi (starogrčki. - boja i tijelo) - nukleoproteinske strukture u jezgru eukariotske ćelije (stanica koja sadrži nukleus), koje postaju lako uočljive u određenim fazama staničnog ciklusa (tokom mitoze ili mejoze). Hromozomi su visok stupanj kondenzacije kromatina koji je stalno prisutan u jezgru stanice. Termin je prvobitno predložen da se odnosi na strukture otkrivene u eukariotskim ćelijama, ali u posljednjim desetljećima sve više govore o bakterijskim kromosomima. Kromosomi su koncentrisani na većinu nasljednih informacija.
Hromozomi su predstavljeni parovima u jezgru svake sperme i svakom jajetu. Osoba ima 23 para ili 46 kromosoma, sa jednim parom nazvanim polnim hromozomima, jer određuju pol budućeg organizma. Ženske ćelije nose dva X hromozomadok muške ćelije sadrže jedan X i jedan Y hromozom. Y hromozom je manjih dimenzija i sadrži manje gena na svojoj površini od X hromozoma. Kao rezultat deljenja ćelija u jajnicima, sva jaja sadrže jedan X hromozom, dok deljenje ćelija u testisima dovodi do činjenice da polovina spermatozoida sadrži X hromozom, a druga polovina sadrži Y hromozom. Odnosno, u polovini slučajeva, ljudsko jaje oplođeno je spermatozoidima koji nose X hromozom, au drugoj polovini - spermatozoidima koji nose Y hromozom, tako da polovina djece rođene u populaciji treba da bude muškarac, a polovina žena.
Dominantni gen(dominantni gen) - gen izražen u fenotipu bez obzira na prisustvo u genomu drugog alela ovog gena. Prisustvo koje osigurava manifestaciju određene osobine, bez obzira na to da li je drugi gen istog para dominantan ili recesivan.
Recesijaeksplicitni gen(recesivan- genetske informacije koje mogu biti potisnute izlaganjem dominantni gen. Recesivni gen je u stanju da osigura manifestaciju određenog svojstva samo ako je uparen s odgovarajućim recesivnim genom. Ako je uparen sa dominantnim genom, onda se ne manifestuje, jer ga dominantni gen potiskuje. Prikazane nekretnine recesivni geni, pojavljuju se u fenotipu potomka samo ako oba roditelja imaju recesivni gen.
Kada su oba alela u paru potpuno identična (npr. OO, AA), onda se takav genotip i njegov vlasnik nazivaju homozygous i kada su ti aleli različiti (recimo, AO) - heterozygous. Poznato je da ako homozigotni genotipovi OO i AA predodređuju prvu i drugu krvnu grupu, tada će vlasnici heterozigotnog AO genotipa imati i drugu krvnu grupu. To znači da se u takvoj kombinaciji ispoljava efekat gena A i efekt gena O, tj. Gena A dominirai gen O u odnosu na njega recesivan (riječ "recesivno" znači nestajanje). Dakle, dominantni geni vrše svoje djelovanje iu homozigotnom i heterozigotnom stanju, a recesivni geni se mogu manifestirati samo u homozigotnom stanju i ne daju vanjske manifestacije u heterozigotnim ljudima.
Skup nasljednih faktora tijela (geni) zove se genotip. Kombinacija svih karakteristika i svojstava organizma, koji su rezultat interakcije genotipa i vanjskog okruženja, zove se fenotip. To znači da je fenotip odraz genotipa u kombinaciji sa okolinom. Granice u kojima okruženje utiče na manifestaciju genotipa naziva se reakcijska norma.
Izvori informacija:
Osnove genetike i uzgoja pasaGenetika je izuzetno teška nauka. Postoje mnogi izuzeci od pravila i mnogi su još nepoznati. Međutim, svi uzgajivači pasa treba da pokušaju da shvate najjednostavnije u Mendelovoj genetičkoj teoriji - „dominantni“ ili „recesivni“ gen ili osobina. Ovo je lako razumljivo i može se direktno primijeniti na uzgoj pasa, jer značajan broj ozbiljnih nedostataka u rasi nastaje zbog recesivnih gena.
Recesivni gen se ne može manifestirati u jednoj ili više generacija dok se ne pronađu dva identična recesivna gena iz svakog roditelja. Psi koji imaju samo jedan recesivni gen neće pokazati ovu osobinu, jer manifestacija recesivnog gena će biti maskirana uticajem dominantnog gena uparenog s njim. Takvi psi / nosioci recesivnog gena / mogu biti opasni za pasminu, ako ovaj gen određuje pojavu nepoželjne osobine time što će je prenijeti na svoje potomke i ostat će u pasmini. Ako slučajno spojite dva psa, nosioca takvog gena, oni će dati dio potomstva neželjenim osobinama.
Dominantni gen se uvek jasno i spolja manifestuje odgovarajućom osobinom. Stoga su dominantni geni koji nose nepoželjne osobine znatno manji rizik za odgajivača, jer njihovo prisustvo se uvek manifestuje, čak i ako dominantni gen funkcioniše bez partnera.
Ali, očigledno, da bi se sve zakompliciralo, nisu svi geni apsolutno dominantni ili apsolutno recesivni. Neki geni su dominantniji od drugih i obrnuto.
Uparivanje ne daje uvek odnos u tačnoj saglasnosti sa očekivanim prosečnim rezultatima i da bi se dobio pouzdan rezultat ovog uparivanja potrebno je proizvesti veliko leglo ili veliki broj potomaka u nekoliko legala.
Genetička tabela pokazuje jednostavnu interakciju dominantnih i recesivnih karakteristika prema Mendelu
Legenda: A - dominantni jednorodni gen
a - recesivni gen jednog roditelja
AA - par dominantnih gena, jedan od svakog roditelja
aa - par recesivnih gena, jedan od svakog roditelja
Aa je dominantni gen jednog roditelja i recesivnog drugog, čineći dominantni recesivni par gena
Objašnjenje opcija:
1. Dva roditelja imaju dva dominantna gena, tako da će svi njihovi potomci biti "čisti" po ovoj osobini - AA
2. Jedan roditelj je "čist" dominantni faktor / AA /, drugi samo izgleda "čisto", ali nosi recesivni gen / Aa /. Prema tome, svi potomci će izgledati "čisto" iz recesivne osobine, ali u stvari polovina potomaka će biti zaista čista, imaju dva dominantna gena / AA /, druga polovina će izgledati "čisto", ali svaki od njih će imati recesivni gen / AA / i biće njegov nosilac. Odnos AA: AA - 1: 1
3. Jedan roditelj se spolja manifestuje dominantna osobina, ali je nosilac recesivnog gena / Aa /. Prilikom parenja sa partnerom koji nosi dva recesivna gena i prirodno pokazuje tu osobinu, polovina potomaka će biti slična prvom partneru / Aa /, a druga polovina drugom / aa /. Dakle, čitavo leglo će biti nosilac ovog gena, samo prva polovina - skrivena, druga eksplicitna.
4. Oba roditelja pokazuju dominantnu osobinu, ali svaki nosi recesivni gen koji se ne manifestuje. U leglu će biti jedan dio potomaka čisto "dominantnih", koji imaju dva A / AA /, dva dijela potomaka, koji izvana pokazuju dominantnu osobinu i stoga, ne razlikuje se od prvog, nosiće recesivno svojstvo, imajući Aa, a jedan dio potomaka će pokazati recesivni karakter znak dva recesivna aa gena. To znači da će odnos potomaka koji pokazuju gestu A koja je određena genomom A biti 3: 1, a prava definicija nosilaca recesivnog i dominantnog heenova će biti: AA: Ah: aa - 1: 2: 1
5. U ovom paru, jedan roditelj je “čisti” dominantni / AA /, drugi ima dva recesivna gena / aa /. Sva legla će izgledati kao dominantan roditelj, ali svaki od potomaka će nositi recesivni gen / Aa /
6. Oba roditelja pokazuju recesivni simptom, jer nose dva recesivna gena / aa /. Sva legla ovog para će nositi ovu osobinu u eksplicitnom obliku i proslijediti je svojim potomcima sve dok se ne upare sa partnerom sa dominantnim genom. U ovom slučaju, ishod uparivanja će biti kao u paru od 3 i 5.
Izvor: Knjiga Hillery Harmar "Psi i njihovo uzgajanje"
Da bi se grubo zamislilo šta potomstvo može doći od datog para proizvođača, potrebno je znati osnovna pravila nasleđa i koje su karakteristike recesivne i koje su dominantne.
Evo liste najčešćih osobina identifikovanih recesivnim genima za većinu pasmina:
Snack, nadglas, lagani nos, svetle oči, dužina udova, kratka njuška, male uši, uspravne uši. Mnogi od teških nasljednih defekata su također kontrolirani recesivnim genima: rascjep usne, rascjep nepca, uvijeni rep, gluvoća, kongenitalna kila, zubi, strah od oštrih zvukova, mokrenje kada je uzbuđen, torzija stoljeća, preokret stoljeća, albinizam, katarakte, itd.
Lobanja i oblik glave su naslijeđeni prema principu Nielson-Ele, a geni koji se jedinstveno djeluju nazivaju se polimerni, a sam nasljeđivanje polimera / široka gornja čeljust dominira uskim, a širina baze lubanje je također naslijeđena. Široki oblik lubanje i donje vilice dominira uskim. Ne potpuna dominacija ima pravokutnu glavu iznad jednog klinastog oblika - i potpuna dominacija ima glavu u obliku buldoga iznad normalne.
Visina i težina se nasleđuju posredno.
HH viseće uši dominiraju stojećim hh, ali postoje i srednje opcije - Hh polu-viseće uši
Prisustvo zubaca na zadnjim ekstremitetima dominira njihovim odsustvom.
Ogromnost dominira nad širinom.
Taxonog dominira normalnom strukturom nogu.
Ekstremne opcije - lagane i teške kosti dominiraju u srednjim tipovima.
Glatka kosa je dominantna u odnosu na dugokose.
Ukrasne dlake su dominantne u odnosu na njihovo odsustvo.
Preovlađuju se obrisi kose u odnosu na njihovo odsustvo.
Knjiga Hilari Harmar "Psi i njihovo uzgajanje"
Da biste razumeli značenje uzgoja, morate početi od samog početka - od rođenja novog života. Ćelija je jedinica života i potiče iz druge žive ćelije. Svaka životinja je proizvod spajanja dve roditeljske ćelije u jednoj ćerki.
Kada se rodi novi život, jedna i samo jedna ćelija sperme (ćelija oca) prodire kroz ljusku jajeta (matična ćelija). Ljuska jajeta onda se odmah zgusne i sprečava prodiranje druge sperme. Ali pre sazrevanja spermatozoida, muška ćelija koja sadrži dvostruki set hromozoma je podeljena na dva dela tako da polovina hromozoma (jedan skup) ulazi u jedan deo, a druga polovina (u isti skup) u drugi. Dakle, u svakom spolu muška ćelija-sperma sadrži jedan set hromozoma. Ženska reproduktivna ćelija, jaje, formirana je na sličan način i sadrži isti set hromozoma. Kombinovanjem dve polovine - dve zametne ćelije - ćelije sperme i jajne ćelije - u kojoj svaki hromozom pronalazi "partnera" iz druge ćelije, formira novu ćeliju iz koje se može razviti novo živo biće, noseći hromozome oba roditelja, opet dupli set hromozoma.
Ali nisu svi hromozomi "partneri" u fuziji roditeljskih zametnih ćelija, na primjer, kromosomi koji određuju spol.
Ženska ćelija ima samo jedan tip spolnog hromozoma, koji se naziva X hromozom. Muška zametna ćelija može imati X i Y kromosome, ali ne oboje. Koja od njih će se povezati sa ženskim X hromozomom je stvar slučajnosti. Ako je X hromozom, tada će klica biti ženski "XX". Ako je Y-hromozom, onda će embrion biti muški "XY", što je karakteristično za sve sisare.
U trenutku spajanja dve polovine roditeljskih ćelija počinje novi život. U tako malom kavezu postoji prototip budućeg psa, iz njega se formira krv, nervni sistem, vuna, mišići, kosti, boja očiju zavisi od njega, poda - zapravo, čitavog budućeg psa.
Prva ćelija je podeljena na dve, zatim četiri, osam, šesnaest, itd., I oni se brzo množe. Svaka ćelija ima nukleus, koji je najvažnija komponenta. Sadrži mrežu struktura nalik nitima koje se protežu u niti nazvane hromozomi. Parovi hromozoma (jedan od svakog roditelja) se drže zajedno i nose mnogo ultramikroskopskih jedinica koje se nazivaju geni, i one su uparene.
Gen je jedinica nasleđa i jedina fizička veza u lancu koji povezuje generacije. Geni su raspoređeni tako da svako zauzima posebno mjesto na vlastitom kromosomu. Svaki gen nosi svoje nasledne osobine i kontroliše ispoljavanje različitih fizičkih i mentalnih karakteristika odraslog živog bića. Pas ima 78 hromozoma (39 parova) koji sadrže hiljade gena. Geni ostaju nepromijenjeni iz generacije u generaciju, osim ako se ne dogodi mutacija. Kod kromosomskog nasleđa, uticaj roditelja je isti, osim u slučajevima nasleđivanja povezanog sa seksom. Geni jednog hromozoma nasleđuju se, po pravilu, zajedno. Svaka specifična karakteristika ili osobina živog organizma određena je njenim genomom, od kojih je polovina naslijeđena od jednog od roditelja, pola od drugog. Za razvedene je najvažnije da znaju za one nosače gena za likove koji imaju dominantno-recesivni odnos.
Recesivni geni
Recesivni gen (to jest, određena osobina) NE MOŽE OBJAVITI JEDNU ILI MNOGU GENERACIJU dok se ne susreću dva identična recesivna gena iz svakog od roditelja (nagla manifestacija takve osobine kod potomaka ne treba miješati s mutacijom).
Psi koji imaju samo JEDAN RECESIVNI GEN - determinantu bilo koje osobine - neće pokazati ovu osobinu, jer će efekat recesivnog gena biti maskiran manifestacijom uticaja para DOMINANTNI GEN. Takvi psi (nosioci recesivnog gena) mogu biti opasni za pasminu ako ovaj gen određuje pojavu nepoželjne osobine, jer će biti prenešeni na sopstvenu hranu, a oni će se nastaviti; i gen je tako očuvan u rasi. Ako slučajno ili nepromišljeno uparite dva nosioca takvog gena, oni će dati dio potomstva neželjenim osobinama.
Genetička tabela koja pokazuje jednostavnu interakciju
dominantne i recesivne karakteristike Mendela
Opcija parenja .... Roditelji ............... Potomci
1 ............. ……………………. AA x AA ....... ..... ..... AA AA AA AA
2 …… .......... …… ... …………. AA x Aa ....... ..... ... .AA AAA Aa Aa
3 ……… .......... …… ... ………. Aa x aa …… ...... ... .aa aa aa aa
4 ……… .......... ……… ... ……. Aa x Aa ... ... ..... ... AA Aa Aa aa
5 ………… .......... …… ... …… .... AA x aa …… .....… .aa aa aa aa
6 ………… .......... …… ... …… .... aa x aa …… ...... ... .aa aa aa aa
A - - dominantni gen jednog roditelja
- recesivni gen jednog roditelja
AAA - par dominantnih gena, jedan od svakog roditelja
aa - par recesivnih gena, jedan od svakog roditelja
Aaa je dominantni gen iz jednog roditelja i recesivnog gena iz drugog,
sastojci dominantno recesivnog para gena
Objašnjenje opcija:
1. Oba roditelja imaju dva dominantna gena, tako da će svi njihovi potomci biti “čisti” na toj osnovi - svi AA.
2. Jedan roditelj je "čist" od dominantnog faktora (AA), drugi samo izgleda "čist", ali nosi recesivni gen (AA). Dakle, svi potomci će izgledati "čisto" iz recesivnog svojstva, ali u stvari polovina potomaka će biti zaista "čista", imaju dva dominantna gena (AA), druga polovina će izgledati kao "čista", ali svaki od njih će imati recesivni gen, tj. biće njen nosilac (Aa). Odnos AA: Aa - 1: 1
3. Jedan roditelj izvana pokazuje dominantnu osobinu, ali je nosilac recesivnog gena (Aa). Prilikom parenja sa partnerom koji nosi dva recesivna gena i, naravno, eksterno pokazuje tu osobinu, polovina potomaka će biti slična prvom partneru (Aa), a druga polovina drugom. Dakle, sve leglo će biti nosilac ove osobine, samo će prvi biti skriveni nosioci, a drugi - eksplicitan. Ovo je posebno važno razumjeti u onim slučajevima gdje je osobina određena recesivnim genom posebno nepoželjna za pasminu.
4. Oba roditelja pokazuju dominantnu osobinu, ali svaki nosi recesivni, ne-manifestni gen. U leglu će biti jedan deo potomaka čisto "dominantnog", koji ima dva A (AA); dva dijela potomaka, koji izvana pokazuju dominantnu osobinu i stoga se ne razlikuju od prvog, također će nositi recesivno svojstvo, imajući Aa; jedan dio potomaka će pokazati recesivno svojstvo, s dva recesivna gena (aa). To znači da će odnos potomaka koji pokazuju genom A, biti 3: 1, a prava raspodela nosilaca recesivnih i dominantnih gena će biti sledeća: AA: A a: a a - 1: 2: 1.
5. U ovom paru - jedan roditelj je “čist” dominantan (AA), drugi ima dva recesivna gena (aa). Sva legla će izgledati kao dominantan roditelj, ali svaki od potomaka će biti nosilac recesivnog gena (Aa), i, shodno tome, osobina koju on određuje.
6. Oba roditelja pokazuju recesivno svojstvo, jer svaki nosi dva recesivna gena (aa). Sva legla ovog para će takođe nositi ovu osobinu i prenositi je dalje na potomke eksplicitno sve dok se ne upare sa partnerom koji nosi dominantni gen. U ovom slučaju, ishod uparivanja će biti kao u paru od 3 i 5.
Očekivani omjer cijepanja potomstva na određenoj osnovi približno je opravdan sa leglom od najmanje 16 štenaca. Za leglo uobičajene veličine - 6-8 štenaca - može se govoriti samo o većoj ili manjoj vjerovatnoći ispoljavanja svojstva, određenog recesivnim genom, za potomke određenog para proizvođača sa poznat genotip.
Dominantni geni
Prisustvo dominantnog gena je uvek jasno i spolja manifestovano odgovarajućom osobinom. Prema tome, dominantni geni koji nose nepoželjnu osobinu predstavljaju mnogo manji rizik za odgajivača od recesivnih, jer se njihovo prisustvo uvijek manifestuje, čak i ako dominantni gen „djeluje“ bez partnera (tj. Aa).
Ali, očigledno, da bi se zakomplicirale stvari, nisu svi geni apsolutno dominantni ili recesivni. Drugim riječima, neki su dominantniji od drugih i obrnuto. Na primjer, neki od faktora koji određuju boju dlake mogu biti dominantni, ali se ipak ne pojavljuju izvana ako ih ne podržavaju drugi geni, ponekad čak i recesivni.
Uparivanje ne daje uvek odnos u tačnoj saglasnosti sa očekivanim prosečnim rezultatima i da bi se dobio pouzdan rezultat ovog uparivanja potrebno je napraviti veliko leglo ili veliki broj potomaka u nekoliko legala.
Neki spoljni znaci mogu biti "dominantni" u nekim stijenama i "recesivni" u drugima. Druge osobine mogu biti posljedica višestrukih gena ili polu-gena koji nisu puke dominante ili Mendelova recesivna. Kao rezultat toga, genetika postaje suviše složena da bi je razumeli prosječni uzgajivač pasa!
Mutacija
Mutacija - nagla promjena u genu. Ona se manifestuje u prvoj generaciji potomaka, ako je mutantni gen dominantan. Ali recesivni mutantni gen može se tajno naslijediti nekoliko generacija, sve dok se dva nosača takvog gena ne odaberu u roditeljskom paru. Tek tada će se pojaviti potomak koji pokazuje rezultat mutacije ovog gena.
Mnoge vanjske promjene uzrokovane su mutacijama. Klasičan primjer ovoga su kvadratne pasmine, kao što su rani mastifi prije nekoliko stotina godina, i sve rase sa skraćenim licem, kao što su pekinezi, mops, buldozi. Pasmine kao što su basovi, pekinezi i jazavčari pate od nasljedno fiksne mutacije koja uzrokuje deformaciju, poznatu kao akondroplazija (abnormalni razvoj tubularnih kostiju ekstremiteta prije rođenja, izražen u smanjenju njihove duljine).
Mutacije su prirodne, ali mogu biti i umjetno uzrokovane, na primjer, ionizirajućim zračenjem (zračenjem). Lijekovi i otrovi mogu biti drugi uzrok i obično uzrokuju štetne mutacije. Uticaj okoline može uticati i na učestalost mutacija. Zanimljivo je da su mutacije nasleđene, tj. Da se uvek reprodukuju, tako da se nove karakteristike ili znaci mogu stalno pojavljivati.
Lethal genes
To su geni koji uzrokuju smrt organizma prije nego što dođe do puberteta. Smrtonosni geni su recesivni, evo nekoliko primera ispoljavanja njihovog uticaja: "rascep usne i rascjep nepca" - defekt u razvoju gornje vilice, hemofilija - nedostatak sposobnosti zgrušavanja krvi, "apsorpcija plodova" u spolno prosperitetnoj kuji, itd.
Polu-smrtonosni geni, na primer, geni koji određuju bilateralni kriptorhizam, postaju smrtonosni za rasu kao rezultat njenog izumiranja. Štenci sa "vučjim ustima", ako nisu operisani, ne mogu da sišu i stoga umiru. Sivo-plava boja sa crnim pjegama povezana je sa polu-smrtonosnim genom i, ako ga naslijedi potomak oba roditelja, onda ovaj potomak može biti slijep, gluh ili bezuspješan. Iz tog razloga, dva psa ove boje nikada se ne pare. Praktično bi bilo najbolje da ovu boju smatramo diskvalifikujućom u svim pasminama.
Razumni uzgoj pasa provodi se kombiniranjem dva glavna tipa uzgoja - INBRIDING i LYNBRIDING, ali s vremena na vrijeme je potrebna i razumna AUTOCROSSING. U prosjeku, potomstvo u svim vrstama uzgoja će biti gotovo jednako, ali gdje je korišćeno inbreeding, posebno u prekomjernoj mjeri, potomstvo će biti ili znatno bolje ili mnogo gore.
Takođe treba imati na umu da čak i najistaknutiji pas pas neće biti u genetskom smislu jednako pogodan za sve kuje.
Inbreeding
Inbreeding je sparivanje bliskih rođaka, na primjer, otac sa kćerkom, majka sa sinom, brat sa sestrom itd. Inbreeding nikada ne treba koristiti ako ne postoji izvjesnost da je stoka apsolutno zdrava, i fizički i mentalno. Ovim načinom uzgoja je strogo obavezno odbacivanje svih uzgojnih materijala koji ne zadovoljavaju standarde. Ako najgori štenci nisu uništeni, oni ne bi trebali proizvoditi pedigree i, naravno, ne bi smjeli dobiti potomstvo od njih.
Neki od najboljih svetskih stoke konja, krava, svinja, pasa i drugih domaćih životinja dobijeni su inbridingom. Ali, primjenjujući inbriding bez dovoljno znanja, moguće je uništiti rasu u nekoliko generacija. Možda će, u jednoj generaciji i, naravno, u dva inbreedinga, pokazati sve mane koje su genetski inkorporirane u pasminu. Treba još jednom naglasiti da je eliminacija svih neuspješnih potomaka iz daljnjeg oplemenjivačkog rada veoma važna tokom inbridinga.
Uzgajivači pasa koji ne znaju pse navedene u pedigreu, najmanje četiri generacije prije, neprikladno je pokušati koristiti inbriding. Rizični eksperimenti su veoma skupi i mogu donijeti veliko razočarenje. Dešava se da inbreeding otkriva nedostatke predaka razdvojenih nekoliko generacija.
Linebreeding
Linebreeding je kao inbriding - rođaci se takođe pare, ali njihov zajednički predak se nalazi samo u trećoj - četvrtoj generaciji. Naravno, što je odnos udaljeniji, to će se više razlikovati vrsta potomaka. Stoga je za prosječnog razvedenog muškarca pametnije zaštititi se i koristiti stroga uzgoj za vaše pse. Kod uzgojnih linija, kao i kod inbridinga, veoma je važno da se iz uzgojnog rada eliminišu svi psi koji ne zadovoljavaju standard.
Outcrossing
Outcrossing je uparivanje nepovezanih proizvođača. Primenjuje se kada je potrebno uvesti neke specifične karakteristike iz druge linije, ili ispraviti nedostatak koji je prodro u liniju u delovima fizičkih ili mentalnih karakteristika.
Ponekad veoma razrađeni parovi daju razočaravajuće pasmine, a on se, konačno, isprepleten, pretvara u potpuni outcrossing koji očekuje od bilo koje mješavine štenaca, ali odjednom pas i kuja iznenađujuće odgovaraju jedni drugima i daju izvanredne lijepe štence. Dakle, jedan potpuni outcrossing može ispraviti nedostatak, ali budite oprezni - potomci takvih križa su rijetko izvanredni pedigree mužjaci i kučke i sami ne proizvode odlične potomke, osim ako nisu bili upareni, tj. vlastitu krvnu liniju koja posjeduje dominantne gene za željene karakteristike.
Ali jednostavno kopiranje uspješnih uzgojnih shema neće donijeti očekivani uspjeh odgajivaču. Ponekad neznanje razvoda o kvalitetu predaka njegovih pasa za uzgoj donosi toliko štete rasi da do vremena kada shvati obim štete, često je prekasno da bilo šta uradi. Stoga je uzgajivač pasa, prije uzgoja pasa, dužan naučiti što više o izabranoj pasmini, o nedostacima koji su najčešći u njemu, o najboljim i najgorim precima uzgojnih pasa s kojima će se razmnožavati.
Najčešći uzrok nedostataka i odstupanja od norme su recesivni geni. Sa približnom tačnošću može se uzeti u obzir:
ako potomak pokazuje karakteristiku koju nijedan od roditelja nije imao,
ovu karakteristiku određuje recesivni gen
Na primer, ako oba roditelja imaju tamne oči ili vrhove nosa, a štene ima svetlost, onda su oba roditelja imala pretka sa recesivnim genom koji određuje takvu osobinu i oni su naslijedili taj gen. Prilikom uparivanja ovih pasa, geni su se spojili - kao rezultat toga, tamnooki roditelji dali su potomke svetlih očiju. Ako se ovo potomstvo pari sa sličnim psom sa svetlim očima, imajući tamnooke roditelje, ni jedan od njihovih štenaca neće imati tamne oči. Ovaj princip se odnosi i na defekte koji su definisani jednostavnim recesivnim genom, kao što je rascjep nepca ili gluvoća. Iz ovoga je jasno kako se nedostatak, određen recesivnim genom, lako može sačuvati i proširiti u rasi, posebno ako je nosač popularan pas, koji u svom životu može proizvesti nekoliko stotina potomaka. Čak i ako nosi samo jedan recesivni gen koji identificira ozbiljnu manu, jasno je, a da sam ne pokaže ovaj nedostatak, on može brzo proširiti ovaj nedostatak u pasmini. Do vremena kada bude otkriveno, biće nemoguće ništa popraviti, pogotovo ako su potomci ovog psa široko korišteni.
Kada su uzgojni psi skloni ekstremnim karakteristikama, pasmina se gotovo sigurno pogoršava. Psi onih pasmina koji, uprkos ljudskoj intervenciji, ostaju srodni svojim divljim precima, ne pate mnogo od raznih defekata koji su fiksirani u drugim rasama kao što su osobenost. Ako su uzgajivači pasa bili svjesni patnji i patnji koje su izazivali svojim psima, a da ne spominjemo da veterinarske službe ispravljaju karakteristike egzotičnih pasmina, tada bi već odlučile da bi bilo pametnije promijeniti standarde. Uzgajivači pasa lako prestaju razmišljati kada su u pitanju njihovi psi, iako su velikodušni s pažnjom na njih, ali brzo postaju slijepi i često previđaju tragedije koje nastaju. Možemo dati samo neke primjere, puna lista bi bila preduga:
Uvijek su teška porođaja kada lubanje i otvori zdjelice ne odgovaraju po veličini jedni drugima (na primjer, kod Boston terijera i francuskih buldoga);
- skraćeni udovi menjaju položaj ilijačnih kostiju karlice, dok je zdjelična rupa preniska, a ako psi ove pasmine imaju opušten trbuh, porod će biti težak (npr. škotski terijeri);
- preduga leđa i bedara daju dodatno opterećenje kičmi, što se odražava u bolesti intervertebralne hrskavice (dachshunds, bassetts);
- prekratke njuške otežavaju disanje, ove pasmine vrlo često, barem češće od drugih, rađaju štence sa vučjim ustima (pekinezi);
- pretjerana slobodna koža formira duboke nabore, gdje se često javlja osip od pelena, kožni kapci zaustavljaju zaštitu očiju, gdje se javlja uporni kronični konjunktivitis;
- previše uski kanal i uzgoj vune stvaraju uslove za uporne bolesti uha (lisne terijere, pudle) ... i mnoge, mnoge druge ...
Nažalost, prirodi nije dozvoljeno da odigra svoju ulogu u uzgoju pasa, svi slabi štenci se spašavaju, hrane, a potom dobijaju potomstvo od njih, dok bi u prirodi morali da umru. Zakon prirode "preživljava najsposobnije za život" ne može se prekršiti nekažnjeno. Dobro je što su mnogi psi sa naslednim defektima neplodni ili manje plodni, tako da najstrašniji nedostaci ne ostaju u rasi, kao što bi mogli uz pomoć "dobrih" uzgajivača pasa.
Kod pasa, kao i kod drugih životinja, uvek je u redu da se najbolje upare sa najboljima, samo onda se uzgajivač može nadati najboljem rezultatu. Postoje mnoge karakteristike i karakteristike koje se ne određuju jednim parom gena, već velikim brojem gena. U hrtovima, na primjer, nemoguće je predvidjeti brzinu trčanja, budući da je određena kombinacijom velikog broja nasljednih faktora, tj. recesivni simptomi za brzinu vožnje. Međutim, poznato je da je u nekim porodicama ili linijama brzina veća, tako da će ponovno spajanje najboljih sa najboljima za dugo vremena sigurno dati najbolje.
Posmatranjem uzgajivača uskoro će se utvrditi koji su geni dominantni. Ali, po mom mišljenju, mnogo je važnije za vlasnika psa da zna koji su geni recesivni, jer oni zahtijevaju najveću pažnju prilikom uzgoja. Nažalost, nisu sve osobine jednako definisane za sve rase dominantnim ili recesivnim genima. Ovo posebno važi za boju dlake.
Možda je vredno ponoviti da ako štene ima znake koji nisu svojstveni bilo kojem od njegovih roditelja, onda su ti znaci kontrolisani recesivnim genima.
Ovdje je korisno dati listu najčešćih karakteristika određenih recesivnim genima za većinu pasmina:
Snack
Nedokus
Light nose
Svetle oči
Dužina udova (za većinu pasmina)
Kratka njuška
Male uši
Uspravne uši (za većinu pasmina)
Dugi kaput (recesivan za glatki sloj)
Glatka (kratka) vuna (recesivna za dugokose)
Mekana vuna (recesivna za tvrdo kosu) \\ t
Ravna vuna (recesivna za kovrčavu) \\ t
Mnogi od teških nasljednih defekata su također kontrolirani genima koji su recesivni za većinu pasmina, na primjer:
rascjep usne, rascjep nepca, uvijeni rep, gluvoća, kongenitalna hernija, zubi, strah od oštrih zvukova, mokrenje kada se uznemire, torzija stoljeća, preokret kapka, albinizam, kamenje mokraćne bešike, hemofilija, zadebljanje i omatanje usana, katarakta i pr.
Boja i pigmentacija vune za mnoge pasmine čvrsto su fiksirane zahtjevima standarda. Ovi karakteri su jedan od najtežih sa stanovišta nasleđivanja, jer kontrolni geni mogu biti dominantni za jednu rasu i recesivnu za drugu, dok kontrolišu istu boju. Što se tiče pigmentacije, sve rase imaju smeđi ili lagani nos, svijetle oči, opće slabljenje pigmentacije određeno je recesivnim genima.
Ako je uzgoj potreban za vođenje po boji, potrebno je napraviti listu svih recesivnih boja, poznatih u ovoj rasi. Za one koji su zaista zainteresovani za uzgoj po boji, snažno preporučujem knjigu Clarencea Litta "Nasleđivanje boja vune u pasa".
Generalizacija
Jedna od najvećih tragedija koja podrazumijeva vlasnika psa je tzv. Ovaj pojam znači da uzgajivač pasa ne vidi nedostatke pasa koji ga uzgajaju, ali je vrlo dobar u sagledavanju nedostataka stoke drugih rasadnika. Međutim, postoji još jedan, možda još veći, defekt kod uzgajivača pasa, takozvana "slepilo za rasu". To se dešava kada nedostatak ne samo puzi, već je i ukorijenjen u pasmini, a stručnjaci u prstenu ga propuste, jer ima vrlo malo pasa koji nemaju takvu manu. Na kraju, nedostatak postaje prihvaćen za pasminu. Tako je, na primer, bilo dislokacija zgloba kolena.
Iskrenost uzgajivača pasa u prepoznavanju grešaka, u njihovoj otvorenoj raspravi je veoma značajna, ali može biti glavna odgovornost za uzgoj najboljih pasa u stvari leži na stručnjacima izložbi. Ako nikada ne dovedu prvog psa do ozbiljnog nedostatka, uzgajivači pasa očito neće potrošiti novac i izložiti takve pse, jer će shvatiti da pas sa ovim nedostacima nema šanse na predstavi. Međutim, sve dok se uzgajivačima pasa dozvoljava da pobijede sa nezdravim psima, onda, uzimajući u obzir ljudsku prirodu, takvi psi ne samo da će i dalje biti izloženi, već će se koristiti i za uzgoj. Neki stručnjaci sa zadovoljstvom pišu u svojim izvještajima da je određeni pas, ili čak i svi psi određene pasmine, koje su ispitivali, izuzetno zdravi, dok je fizički većina njih bila izuzetno nezdrava.
To je loša usluga za pasminu, ali na tako laskav način nećete potrošiti ozbiljnog uzgajivača koji strogo poštuje da pasmina nije "puštena" u greške, posebno one koje narušavaju zdravlje i ugrožavaju samu egzistenciju psa, kao i katarakte, volvuluse, sumrak. i displazije zglobova.
Sada ima previše pasa, a možda i manje i da bolje, ali nažalost, "igra pasa" je uvijek bila prilično privlačna političarima iz uzgoja pasa. Međutim, tokom vremena, uprkos nevažnim stručnjacima i neznanju uzgajivača pasa, većina pasmina se poboljšala i nastavlja da se poboljšava, posebno što se tiče izgled. Ovakav zaključak može se napraviti upoređivanjem današnjih predstavnika pasmina sa fotografijama snimljenim prije 50 godina.
Nažalost, postoje izuzetno veliki broj uzgajivača pasa koji "ne vide" psa i stoga su apsolutno nesposobni primijetiti nedostatke i, posljedično, njihove zasluge. Čovek je zaista rođen sa prirodnom sposobnošću da vidi životinju, onda može da izabere dobrog psa u bilo kojoj rasi. Stručnjaci se takođe rađaju, ne prave, iako se znanje, naravno, može akumulirati.
Najteže je odrediti najboljeg psa kada morate izabrati veliki broj loših ili srednjih pasa. U svakom pogledu, veličanstveni uzorak sa jednom primjetnom manom bolji je od prosječnog psa, koji ima i vrlo dobre i vrlo loše osobine. Očigledno je da je lakše sa daljim uzgajanjem upotreba potomaka ovih pasa osloboditi se jednog nedostatka, a ne pokušati uvesti u pasminu istovremeno mnoge dobre osobine.
Recesivni geni
RECESIVNI GEN (tj. Određena osobina) NE MOŽE IZRAŽAVATI U JEDNOM ILI MNOGU GENERACIJA dok se ne susreću dva identična recesivna gena iz svakog od roditelja (nagla manifestacija ove osobine kod potomaka ne treba miješati s mutacijom).
Psi koji imaju samo jedan RECESIVNI GEN - determinantu bilo koje osobine - neće pokazati ovu osobinu, jer će efekat recesivnog gena biti maskiran manifestacijom uticaja DOMINANTNOG GENA uparen s njim. Ovakvi psi (nosioci recesivnog gena) mogu biti opasni za pasminu, ako taj gen određuje pojavu neželjene osobine, jer će se prenijeti na vlastito poreklo, a oni dalje i tako će ostati u pasmini. Ako slučajno ili nepromišljeno uparite dva nosioca takvog gena, oni će dati dio potomstva neželjenim osobinama.
Genetička tabela pokazuje jednostavnu interakciju dominantnih i recesivnih karakteristika prema Mendelu
Legenda:
A - - dominantni gen jednog roditelja
I -? - recesivni gen jednog roditelja
AA - par dominantnih gena, jedan od svakog roditelja
aa -? ? - par recesivnih gena, jedan po jedan od roditelja
Ah -? - dominantni gen iz jednog roditelja i recesivnog gena iz drugog, čineći dominantan - recesivni par gena.
Objašnjenje opcija:
1. Oba roditelja imaju dva dominantna gena, tako da će svi njihovi potomci biti “čisti” na toj osnovi - svi su AA.
2. Jedan roditelj je “čist” u dominantnom faktoru (AA), drugi samo izgleda “čist”, ali nosi recesivni gen (Aa). Dakle, svi potomci će izgledati "čisto" iz recesivnog svojstva, ali u stvari polovina potomaka će biti zaista "čista", imaju dva dominantna gena (AA), druga polovina će izgledati "čisto", ali svaki od njih će imati recesivni gen, tj. će biti njen nosilac (Aa). Odnos AA: Aa - 1: 1.
3. Jedan roditelj izvana pokazuje dominantnu osobinu, ali je nosilac recesivnog gena (Aa). Prilikom parenja sa partnerom koji nosi dva recesivna gena i, naravno, eksterno pokazuje tu osobinu, polovina potomaka će biti slična prvom partneru (Aa), a drugom drugom. Dakle, sve leglo će biti nosilac ove osobine, samo će prvi biti skriveni nosioci, a drugi - eksplicitan. Ovo je posebno važno razumjeti u onim slučajevima gdje je osobina određena recesivnim genom posebno nepoželjna za pasminu.
4. Oba roditelja pokazuju dominantnu osobinu, ali svaki nosi recesivni, ne-manifestni gen. U leglu će biti jedan deo potomaka čisto "dominantnog", koji ima dva A (AA); dva dijela potomaka, koji izvana pokazuju dominantnu osobinu i stoga, ne razlikuju se od prvog, nose recesivno svojstvo, imajući Aa; jedan dio potomaka će pokazati recesivno svojstvo, s dva recesivna gena (aa). To znači da će odnos potomaka koji pokazuju gestu A određenu genomom A biti 3: 1, a prava raspodela nosilaca recesivnih i dominantnih gena će biti sledeća: AA: Ah: aa - 1: 2: 1.
5. U ovom paru - jedan roditelj je “čist” dominantan (AA), drugi ima dva recesivna gena (aa). Celo leglo ovog para će takođe nositi ovu osobinu, i proslediti je dalje potomcima u eksplicitnom obliku sve dok se ne upare sa partnerom koji nosi dominantni gen. U ovom slučaju, ishod uparivanja će biti kao u paru od 3 i 5.
6. Oba roditelja pokazuju recesivno svojstvo, jer svaki nosi dva recesivna gena (aa). Celo leglo ovog para će takođe nositi ovu osobinu, i proslediti je dalje potomcima u eksplicitnom obliku sve dok se ne upare sa partnerom koji nosi dominantni gen. U ovom slučaju, ishod uparivanja će biti kao u paru od 3 i 5.
Očekivani omjer cijepanja potomstva na određenoj osnovi približno je opravdan sa leglom od najmanje 16 štenaca. Za leglo uobičajene veličine - 6-8 štenaca - može se govoriti samo o većoj ili manjoj vjerovatnoći ispoljavanja svojstva, određenog recesivnim genom, za potomke određenog para proizvođača s poznatim genotipom.
Iz knjige Tajne ljudskog nasljeđa autor Afonkin Sergej YuryevichDominantni i recesivni geni Zamislite dva homologna hromozoma. Jedan od njih je majčinski, a drugi očinski. Kopije gena lociranih na istim DNK segmentima takvih hromozoma nazivaju se alelni ili jednostavno aleli (grčki alios je drugi). Ove kopije
Iz knjige Tajne ljudskog nasljeđa autor Afonkin Sergej YuryevichGeni starenja Starost je gadna stvar, a svake godine se pogoršava. Diane Cooper Ljudi ne žele da žive večno. Ljudi jednostavno ne žele da umru. Stanislav Lem - Postoje besmrtne ćelije - Programirana smrt - Zakretanje starenja - Bolesti
Iz knjige Evolucija čovjeka. Knjiga 2. Majmuni, neuroni i duša autorGeni i ponašanje Činjenice dobijene od neurobiologa govore o materijalnoj, neurološkoj prirodi psihe. Ali da bi se duša u svim svojim manifestacijama mogla razvijati, ovo, strogo govoreći, još uvijek nije dovoljno. Nisu svi znaci mogu evoluirati, već samo
Iz knjige Evolucija čovjeka. Knjiga 1. Majmuni, kosti i geni autor Markov Alexander VladimirovichNovi geni? Aktivnost gena može da se promeni tokom evolucije ne samo promenom mesta vezivanja TF, radom TF-a ili regulatornih RNK, već i kao rezultat dupliciranja gena. Dok su ostale stvari jednake, dva identična gena će proizvesti više proizvoda (tj. Informacije
autor Harmar Hillery Iz knjige Uzgoj pasa autor Harmar Hillery autor Harmar HilleryRecesivni geni RECESIVNI GEN (tj. Određena osobina) NE MOGU BITI ODGOVARANI OD JEDNOG ILI MNOGA GENERACIJA dok se ne nađu dva identična recesivna gena iz svakog od roditelja (nagla manifestacija ove osobine kod potomaka ne treba miješati sa
Iz knjige Psi i njihov uzgoj [Uzgoj pasa] autor Harmar HilleryDominantni geni Prisustvo dominantnog gena je uvek jasno i eksterno manifestovano odgovarajućom osobinom. Prema tome, dominantni geni koji nose nepoželjnu osobinu predstavljaju mnogo manji rizik za odgajivača od recesivnih, jer je njihovo prisustvo
Iz knjige Psi i njihov uzgoj [Uzgoj pasa] autor Harmar HillerySmrtonosni geni Ovo su geni koji uzrokuju smrt organizma prije nego što dosegne pubertet. Smrtonosni geni su recesivni, evo nekoliko primera ispoljavanja njihovog uticaja: "rascep usne i rascjep nepca" je defekt u razvoju gornje vilice, hemofilija je nedostatak krvi
Iz knjige Neandertalci [Istorija frustriranog čovječanstva] autor Vishnyatsky Leonid Borisovich autor Kandel Eric Richard Iz knjige U potrazi za memorijom [Pojava nove nauke o ljudskoj psihi] autor Kandel Eric Richard autor Hengstschleger MarkusGeni sportista Žene ga vole zbog njegovog izgleda, a muškarci vole njegovu sposobnost da zabija lopte. Žene se dive njegovoj frizuri i zavodljivom osmehu, muškarci ga poštuju zbog bacanja sa boka. Radi se o dvoje ljudi. Nakon svega što smo naučili o genetici
Iz knjige Moć gena [lijepa kao Monroe, pametna kao Einstein] autor Hengstschleger MarkusGeni Mocarta O detinjstvu Mozarta ne postoje toliko tačne informacije, mnogo više - pretpostavke. Kako se ovaj mali genij dogodio? Sestra Nannerl, pet godina starija, htjela je znati više o svom mlađem bratu. Kakav je bio u ranom detinjstvu? U
Iz knjige Moć gena [lijepa kao Monroe, pametna kao Einstein] autor Hengstschleger MarkusGENIJI I sada sasvim drugo pitanje: zašto je to tako zanimljivo? Šta možete saznati ispitivanjem lobanje Mozarta? Pa, recimo da je bio meren i meren. Prema rezultatima analize kostiju, možete saznati šta je vlasnik ove lobanje jeo. I to je već učinjeno. Ova lobanja
Iz knjige Unutarnja riba [Povijest ljudskog tijela od antičkih vremena do danas] autor Shubin NilGeni Otkriće Arendta dovodi nas do još jednog pitanja. Jedno je da oči različitih životinja imaju zajedničke delove, ali kako se desilo da su različite oči kao crvi, muhe i miševi međusobno povezane? Da odgovorimo na ovo pitanje, obratimo se
Opći podaci
Rezultati interakcije gena dva roditeljska genoma u zigoti i višestaničnog organizma koji se razvijaju iz njega manifestuju se u osobinama koje kontroliraju, koje se donekle prenose iz generacije u generaciju. Takvo nasleđivanje zavisi od mnogih razloga.
Dugo vremena u klasičnoj genetici, smatralo se da je doprinos oba roditelja genomu potomstva bio približno isti za materinsku i očevu liniju porodičnih rodova. Na osnovu toga formulisali su jedno od prvih pravila nasleđivanja - ekvivalentnost i povezanost funkcija dva alela različitog porekla istih gena (ili ekvivalentnost recipročnih ukrštanja.Identifikovane su dvije varijante i niz tipova nasljeđivanja, uzimajući u obzir broj gena, njihovo porijeklo (majčinsko ili očinsko), lokalizaciju autosoma i spolnih hromozoma u alelnim ili nealelnim lokusima, prirodu manifestacije (dominacija ili recesija), te značajke (mehanizme) interakcije između gena.
Međutim, u drugoj polovini dvadesetog stoljeća utvrđeno: u mnogim slučajevima doprinos jednog roditelja se značajno razlikuje od doprinosa drugog roditelja. Pokazalo se da funkcije roditeljskih gena mogu varirati kroz cijelu ontogenezu sve do diferencijalne obustave majčinskih ili očinskih alela. Osnova ovog fenomena je epigenomski proces ili obeležavanje lokusa hromozoma jednog od roditelja, što dovodi do isključenja ekspresije alela koji se nalaze u njima. Ovaj fenomen se naziva imprinting.Pojam je prvi put korišten u drugoj deceniji 20. stoljeća. Austrijski zoolog Konrad Lorenz. Gledajući ponašanje pačića koji su se samo izlegli iz ljuske, primijetio je: oni traže svoju majku patku. Ako vlasnik ili domaćica patke, ili pas ili mačka koja prođe, dođe u njihov pogled, pače sledi čitav svoj budući život kao njihova majka.
Dakle, govorimo o formiranju prije rođenja brojnih tjelesnih funkcija utisnutih u genom (“genomska memorija”).
Kasnije je pojam "imprinting" počeo da se koristi od strane neuropatologa. Vjeruje se da je tiskanje posljedica niza urođenih fenomena kod novorođenčadi, kao što je potraga za bradavicom majčinih žlezda ili osnovnih funkcija (gutanje, disanje, žvakanje, cirkulacija krvi, probava i sisanje), koje su nastale u intrauterinom životu.
Tada je ovaj termin počeo da koristi genetika da objasni neobično ponašanje seksualnih hromozoma (eliminacija hromozoma paternalnog porekla) kod insekata roda Sciara Coprophila (G. Crouse, 1960). Pokazano je da su paternalni X kromosomi nekako označeni (utisnuti) prije spajanja gameta u skladu sa njihovim roditeljskim porijeklom.
Trenutno je ubedljivo dokazano opšte biološko značenje imprintinga kao epigenomskog procesa povezanog sa regulatornim (funkcionalnim) promjenama u aktivnosti genoma, a koje nije povezano sa strukturalnim oštećenjem (čl. 28).
Kod ljudi, kao rezultat imprintinga, na primer, izražava se materinski gen (alel), dok je alel na očinskoj osnovi blokiran ili obrnuto. Shodno tome, pojedinac ima samo monoalelni izraz ili neekvivalentan doprinos genomu jednog od njegovih roditelja, što znači da postoji odstupanje od Mendelovih zakona o nasleđivanju zasnovanih na dialelnom modelu (vidi Poglavlje 2).
Prije razmatranja takvih odstupanja, osvrnimo se na obrasce nasljeđivanja gena i osobina, koje je najprije opisao G. Mendel na temelju rezultata njegovih eksperimenata na križanju sjemena vrtnog graška. U budućnosti, ovi obrasci se zovu zakoni nasleđivanja i šire se na ljude.
Zakoni nasleđivanja
U modernom izdanju, zakoni nasleđa su formulisani na sledeći način.
Prvi zakon- zakon dominacije (ili uniformnosti) osobine u potomcima prve generacije. U prvoj generaciji, potomak manifestuje djelovanje dominantnog gena (dominantan
simptom), ali se efekat recesivnog gena ne manifestuje (recesivno svojstvo). U narednim generacijama, potomak manifestuje djelovanje i dominantnog i recesivnog gena.
Drugi zakon- Zakon o razdvajanju gena u potomstvu ili zakonu "čistoće gameta". U potomstvu se u gametama javlja alternativna podjela (distribucija) gena: jedna polovina gameta nosi dominantne gene (A), a druga polovina nosi recesivne gene (a). Oba tipa gena su prisutna u somatskim ćelijama, ne miješajući se i ne zamjenjujući se.
Treći zakon- zakon nezavisno nasleđivanje nealelni geni ili slučajne kombinacije nasljednih sklonosti kod potomaka. Nasleđivanje dva gena (dva para znakova) se zove dihybrid,više od dva gena (dva para znakova) - polyhybridFormula za takvo nasleđivanje odgovara binomijalnoj seriji: (3 + 1) n, gde je η broj gena (para znakova). Za izvođenje formule koristi se rešetka R. Pennet. Uz njegovu pomoć izračunavaju se genotipovi organizama, njihov broj i zavisnost od tipova gameta koji sadrže dominantne i recesivne gene (Tabela 3).
Tabela 3.Rešetka R. Penneta
NapomenaA, B, A "i B" - dominantni geni; a, b, a "i b" su recesivni geni; A "A, B" B, A "B i B" A - dominantni homozigotni organizmi; a "a, a" b, b "a, b" b - recesivni homozigotni organizmi; A "a, a" b, a "A, a" B, B "a, B" b, b "a i b" B - dominantni heterozigotni organizmi.
Pored tri zakona nasleđivanja, nasleđivanje gena i karaktera određuje glavne odredbe hromozomske teorije nasleđa.
Hromozomska teorija nasljednosti
U drugoj deceniji XX veka. Thomas Hunt Morgan (1866-1945) i njegovi učenici (C. Bridges, G. Meller i A. Stertevant) glavne odredbe kromosomske teorije nasljednosti(sve
Moderna hromozomska teorija nasleđa uključuje sledeće odredbe.
Geni se nalaze u kromosomima; broj gena u hromozomu je proporcionalan dužini hromozoma. Kasnije se ispostavilo: različiti geni su različiti po dužini, tako da se ova proporcija ne poštuje uvijek. Na primer, na hromozomima 5 i 9, geni su identifikovani više nego na najdužim hromozomima 1 i 2.
Geni se nalaze duž dužine kromosoma na linearan način. Nakon toga, identifikovani su mobilni genetski elementi ili „skakački geni“ (transpozoni) koji se kreću duž genoma od jednog kromosoma do drugog, ometajući linearni poredak.
Alelni geni zauzimaju identične lokuse homolognih hromozoma. Od dva homologna hromozoma porekla, jedan je otac, a drugi majka. Lokusi su međusobno identični. Oni sadrže alele istog gena ili alelnih gena. Svaka od dva alela istog gena je očinska i majčinska kopija; normalno, po pravilu, to je dialelni model organizacione strukture gena (vidi sliku 12). U slučaju nasljedne patologije, pojedinac može imati samo jedan lokus gena, te će u skladu s tim postojati jedan alel (ili očinski ili majčinski); ovo monoalelno stanje ovog alela je zabeleženo, na primer, kod Shereshevsky-Turner-ovog sindroma (kariotip: 45, XO). Druga patološka varijanta je prisustvo pojedinca istovremeno tri alela i više; to je stanje polilelizma za ovaj alel - na primjer, tri identična alela istog gena tri hromozoma 21 u Downovom sindromu (kariotip: 47, XX, + 21); tri alela od tri hromozoma 13 u Patau sindromu (kariotip: 47, XY + 13) ili prisustvo u kariotipu od 4 do 11 X hromozoma (4-11 alela istog gena - 49, XXXX).
Geni istog hromozoma čine grupu za povezivanje, osiguravajući zajedničko nasleđivanje osobina koje kontrolišu. Treba napomenuti: povezivanje gena sa hromozomom je stalno poremećeno tokom prelaska - proces homologne rekombinacije ili razmena identičnih lokacija (geni i njihovi fragmenti).
između homolognih hromozoma u prvoj podjeli mejoze. Učestalost križanja je direktno proporcionalna udaljenosti između gena. Međusobna razmjena alela paternalnog i majčinskog porijekla javlja se između svih parova homolognih kromosoma, s izuzetkom X i Y kromosoma. Otvaranje prelaza pripada i školi T.H. Morgana Sada razmotrite opcije i tipove nasleđivanja gena i osobina.
OPCIJE I VRSTE GENA I ANKETA O SIGNALIMA
Trenutno postoje tri varijante nasleđivanja gena i karaktera: monogeno i poligensko sa tradicionalnim (klasičnim) nasleđem i ne-klasičnim ili netradicionalnim nasleđem.
Monogeno nasleđivanje se zasniva na prvom i drugom zakonu nasleđa. To podrazumeva nasleđivanje jednog gena (jedan par karaktera) i odnosi se na alelne gene.
Poligensko nasleđe se zasniva na trećem zakonu nasleđa. To podrazumeva nasleđivanje dva gena (para znakova) i više i odnosi se na nealelne gene.
Netradicionalno nasleđivanje je nasleđivanje gena i osobina koje prevazilaze monogene i poligene varijante.
Monogeno nasleđivanje
Monogeno nasleđivanje se često naziva jednostavnim Mendelovim nasleđem.
Na osnovu ideja o dialelnom modelu strukture gena, nasleđivanje mehanizama interakcije između roditeljskog i majčinskog genoma se razmatra odvojeno za svaki alelni i nealelni par.
Vrste monogenog nasljeđivanja
U okviru monogenskog nasljeđivanja, postoje:
Autosomno dominantan tip (dominantni gen nalazi se na jednom od dva autosoma);
Autosomalno recesivni tip (recesivni gen se nalazi na jednom od dva autosoma);
X-vezani dominantni tip (dominantni gen se nalazi na X hromozomu);
X-vezani recesivni tip (recesivni gen se nalazi na X hromozomu);
Y-vezani tip ili holandrično nasleđe (gen se nalazi na Y hromosomu).
Prema katalogu V. McKusicka "Nasljeđivanje Mendelovih osobina kod ljudi" (Internet verzija: online - http: www.ncbi.nlm. Nih.gov/Omim), identificirano je više od 12,5 tisuća takvih fenotipa. Među njima, oko 12 hiljada su fenotipovi koji su nasleđeni (ili verovatno nasleđeni) autosomno dominantnim ili autosomalno recesivno (uključujući 9 hiljada fenotipa sa ustanovljenim tipom nasleđivanja).
Poslednjih godina dobijeni su brojni podaci o nasleđivanju vezanim za X i Y hromozome, na kojima su lokalizovani.
300 i 92 gena.
Na sl. 20 prikazuje mapu X hromozoma i broj dominantnih i recesivnih bolesti povezanih sa svojim lokusima.
Sljedeće bolesti koje se rijetko javljaju treba navesti kao primjere X-vezanih dominantnih fenotipova: rahitis ili hipofosfatemiju otpornu na vitamin D (22,2); sindrom pigmentne inkontinencije, tip I (Xp11.1) i tip II (Xq28).
Primjeri najčešćih bolesti naslijeđenih X-vezanim recesivnim tipom: hemolitička anemija (Xq21.2 ili Xq28), hemofilija A (Xq28.2) i B (Xq27.2), Duchenne-Becker mišićna distrofija (Xp21.2), Lesch sindromi -Nayana (Xq26.2),
point condysplasia od Konradi-Hünermann (Hr 22.2), retinitis pigmentosa (Xp21.2-21.3; Xp22).
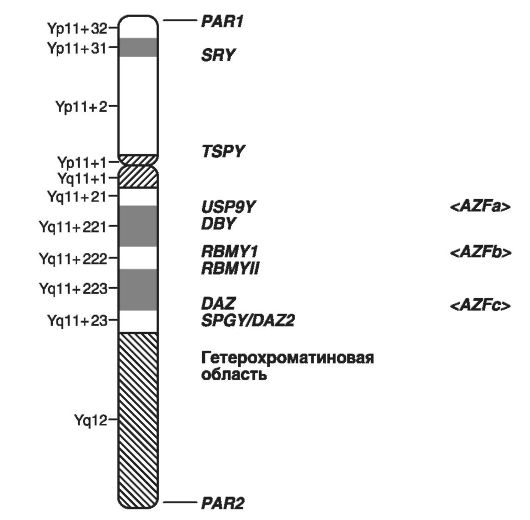
Na sl. Slika 21 prikazuje šematski prikaz lokusa Y-hromozoma, u kojima se nalaze geni koji formiraju muški spol i geni koji uzrokuju mušku neplodnost.
Kršenja seksualne diferencijacije, oblici muške neplodnosti u obliku azoospermije (Yp - faktor 2; Yq11 - faktor 1), gonoblastoma, itd. Treba navesti kao primjere bolesti povezanih s Y kromosomom, ukupno geni odgovorni za sedam takvih bolesti su istaknuti u dijagramu. .
Pored toga, monogeno nasleđivanje može biti ne samo vezan za pod (X ili Y hromozom), već i ograničen na pod.
Sl. 20Karta ljudskog X hromozoma (prema multimedijalnom udžbeniku "Biology" Copyright (c), 2007)
Na primer, ćelavost se manifestuje kod muškaraca (dominantan efekat) i gotovo se ne ispoljava kod žena (recesivni efekat).
Kriterijumi za monogeno nasleđivanje
Autosomno dominantan tip:
Bolest se redovno prenosi iz generacije u generaciju bez praznina, tj. u poreklu u vertikalnom pravcu, osim u slučaju de novo mutacija (vidi Poglavlje 5);
Sl. 21.Shematski prikaz Y kromosoma (prema GenBank, 2003)
Rizik da ima bolesno dijete ako je jedan od roditelja bolestan je 50%;
Zdravi pojedinci imaju zdravo potomstvo;
Bolestan pacijent ima jednog od roditelja, osim u slučaju nove mutacije;
Oba pola su pogođena istom frekvencijom. Autosomno recesivni tip:
Roditelji probandnog pacijenta (osobe koja je konsultovala genetskog lekara) su zdravi, ali sličnu bolest ima i rođaci, rođaci i rođaci probanda (njegova braća i sestre), tj. vodoravno u poreklu (u jednoj generaciji);
Bolestan roditelj rađa zdravu djecu;
Rizik obolevanja od bolesnog djeteta je 25% (omjer bolesnih i zdravih osoba je 1: 4);
U slučaju nasljednih brakova između roditelja probandnog pacijenta, u pedigreu se povećava broj bolesnih rođaka.
Dominantni X-vezani tip:
Pacijent proband nužno ima jednog roditelja;
Bolestan otac bolestan je za sve svoje kćeri, a sinovi mu zdravi;
Bolestna majka jednako je vjerovatno da ima bolesnu kćer i bolesnog sina;
Kod zdravih roditelja, sva djeca će biti zdrava;
Ima 2 puta više bolesnih žena nego bolesnih muškaraca. X-recesivni tip:
Bolest je uočena kod muških rođaka pacijentovog probanda na majčinoj liniji;
Sinovi ne nasljeđuju očinsku bolest;
Kod bolesnog oca sve kćeri su zdrave i heterozigotni nosioci gena bolesti oca;
Ako je žena heterozigotni nosilac gena bolesti, onda će polovina njenih sinova biti bolesna i sve kćeri su zdrave; a polovina ćerki će takođe postati heterozigotni nosioci gena bolesti.
Kriterijumi za holandsko nasleđivanje još nisu razvijeni. Poslednjih godina, u ljudi je identifikovano više teških nasleđenih monogenih i poligenih bolesti:
Digestivne bolesti - rak dojke (dvije genokopije: geni VRS1 i VRS2);
Trigenske bolesti - Bardet-Beadle sindrom (3 genokopije: geni BBS1, BBS2 i BBS30);
Kvadgenske - Pentagene bolesti: Alzheimerova bolest (uzrokovana četiri genokopije: PS1, PS2, PS3 i PS4, kao i genom prionskog proteina, vidi poglavlje 29);
Poligene ili multifaktorne bolesti (MDF) uzrokovane "mrežama gena" - arterijska hipertenzija (170 gena, uključujući 17 glavnih gena), bronhijalna astma (oko 20 gena), osteoporoza (13 gena), endometrioza (11 gena), itd.
Poligensko nasleđivanje
Poligensko nasleđe se često naziva multigene ili multifaktorijalno, što znači da se u isto vreme ne nasleđuje ni jedan već nekoliko specifičnih gena koji ispoljavaju svoje
djelovanje u specifičnim uvjetima okoline, u prisustvu izazivačkih vanjskih faktora, po pravilu, pojačava individualno djelovanje gena, čiji je učinak kumulativni (aditivno djelovanje).
Kriterije poligenskog nasljeđivanja (ukupno 5) sistematizirano je 1969. godine K. Carter, a 20 godina kasnije dopunjeno je (još dva) F. Vogelom i A. Motulskim (1989).
Kriterijumi poligenetskog nasleđivanja
Rizik od razvoja multifaktorijske osobine (bolesti) određen je sljedećim faktorima.
Nasleđivanje neke osobine ili bolesti. Što je veća heritabilnost neke osobine ili bolesti (što su za to odgovorni naslijeđeni geni), to je veći rizik od njegovog razvoja kod zdravih rođaka. Nasljeđivanjeznak (bolest) je stepen uticaja na formiranje datog znaka (bolesti) nasljednih faktora u odnosu na faktore okoline. Nasleđivanje se izražava u apsolutnim brojkama (od nula do jedan) ili u procentima koristeći koeficijent h 2 ili K n, koji se izračunava po formuli: K n = G / E x 100%, gde je K n koeficijent nasleđivanja, G je nasledni faktor, E - faktori okoline. U tabeli 4 prikazane su vrijednosti broja koeficijenata nasljeđivanja multifaktorijskih osobina i bolesti.
Tabela 4.Koeficijenti nasleđivanja multifaktorskih znakova i bolesti
Ozbiljnost simptoma ili težina bolesti u probandu. Što je znak jači ili je teže curenje
bolest u bolesnog rođaka, veći rizik od njegovog razvoja kod zdravih rođaka.
Zajedničnost gena u probandu i njegovih rođaka (ili bliski stepen srodstva sa bolesnim rođakom). Što su uobičajeni geni kod pacijenta i njegovih rođaka, veći je rizik od razvoja znaka ili bolesti u potonjem. Na primjer, učestalost populacije psorijaze je 0,75%. Među srodnicima prvog stepena srodstva, učestalost njegovog razvoja je 5,6%, među srodnicima drugog stepena srodstva - 3,0–3,5%, među srodnicima trećeg stepena srodstva - 1,75%, među rođacima četvrtog stepena srodstva - 0,75% .
Retko se javlja seks. Multifaktorski znak ili bolest se češće javlja kod pojedinaca retko pogođenog spola (što se zove kriterij carter efekat).Na primjer, kongenitalna stenoza pilorusa javlja se kod dječaka 2-5 puta češće nego kod djevojaka, tj. u ovom slučaju, ženski spol je retko pogođen seks. Međutim, učestalost ove bolesti u buduće djece djevojčica oboljelih od stenoze pilorusa će dostići 10-20%, dok će u budućnosti djeca dječaka pogođena stenozom pilorusa - samo 2-6%. Drugi primjer je čir na želucu i duodenalni ulkus, koji se po pravilu manifestiraju kod muškaraca i mnogo rjeđe kod žena. Međutim, njegova učestalost kod djece bolesne žene je veća nego kod djece bolesnog muškarca.
Broj bolesnih rođaka. Što je više rođaka u pedigreu koji imaju multifaktorijsku osobinu ili bolest, to je veći rizik njegovog razvoja među potomcima (Tabela 5).
Tabela 5.Rizik razvoja multifaktorske osobine ili bolesti u probandu, u zavisnosti od broja bolesnih srodnika
Ukupni rizik za djecu, ako su njihovi roditelji zdravi, je 5-10%; ako je jedan od roditelja bolestan - 10-20%; ako su oba roditelja bolesna - do 40%;
Dodatni kriteriji
Dvostruki kriterijum. Ako je podudarnost (sličnost) jednozigotnih blizanaca na nekom znaku ili bolesti 4 puta veća od podudarnosti kod dizigotskih blizanaca, onda je ovaj znak ili bolest naslijedila prema poligenskoj varijanti.
Kriterijum segregacije pogođenih braće i sestara u porodicama sa jednim bolesnim ili dva zdrava roditelja. Ako je udio bolesnih braće i sestara u porodicama s jednim bolesnim roditeljem 2,5 puta veći (i više) od udjela bolesnih braće i sestara u porodicama sa dva zdrava roditelja, tada je naročito vjerovatna poligenska varijanta nasljeđivanja. Ako je ovaj odnos manji od 2,5, tada nije isključeno i poligeno nasleđivanje.
Međutim, klasične varijante nasljeđivanja gena i osobina javljaju se samo u manjem dijelu svih slučajeva takvog nasljeđivanja, dok ih je gotovo 2/3 netradicionalne varijante.
Nontraditional nasledstvo
Netradicionalno nasleđe primećeno je kod mnogih naslednih bolesti čoveka. Proteklih godina im je dodijeljeno prethodno slabo proučeno ili općenito nepoznato oboljenje od 6 klasa: akumulacijske bolesti (44 nosologije; vidi poglavlje 21); peroksizomalne bolesti (17 nosologija; vidi poglavlje 26); mitohondrijske bolesti (istaknuto je više od 200 nosologija, pretpostavlja se još 1150; vidi poglavlje 26); imprinting bolesti (istaknuto je 24 nosologija; vidi poglavlje 28); bolesti ekspanzije broja ponavljanja nukleotida (27 nosologija; vidi poglavlje 27); prionske bolesti (9 nosologija; vidi poglavlje 29).
U principu, netradicionalne bolesti nasleđa čine 321 nozološku jedinicu (isključujući 1.150 mitohondrijskih bolesti). Nasleđujući bolesti prve tri klase nasljedstvo majkejer lizosomi, mitohondrije i peroksizomi, kao citoplazmatske strukture somatskih ćelija, nasleđuju se isključivo preko linije majka-ćerka i nikada se ne prenose preko linija majka-sin, otac-sin ili otac-ćerka, što je zbog biološka matrijarhija(vidi poglavlja 5 i 12).
Međutim, u brojnim mitohondrijskim bolestima, pored čisto majčinskog nasleđivanja, može se primetiti monogensko nasleđe mešano sa majčinim. To je povezano sa poremećenom interakcijom između mitohondrijskih (mtDNA) i nuklearnih (DNK) genoma. Posebno, geni koji kodiraju mitohondrijske proteine nalaze se ili u mitohondrijima (geni dva tipa rRNK, 22 tipa tRNA, 13 tipova polipeptida), ili u jezgru ćelija (njih ima oko 1150). Nuklearni geni se transkribuju u jezgru, prevedeni u citoplazmu, a rezultati njihove ekspresije „uvoze“ mitohondrije.
Obično, sve kopije mtDNA su identične jedna drugoj (homoplazija). U slučajevima kada se mutacije dešavaju u mtDNK (njihova učestalost je 10 puta češća nego u nuklearnoj DNK), moguće su dvije vrste mtDNA u jednoj ćeliji (heteroplazija), a zatim se naziva proces propagacije mutantne i normalne mtDNK. replikativna segregacija(vidi poglavlje 2).
U slučaju imprinting bolesti ili epigenomske deaktivacije ekspresije jednog od roditeljskih kromosomskih lokusa (vidi gore), fenotipske manifestacije genskog djelovanja mogu se promijeniti kao rezultat tri razloga: brisanja gena (genetički imprinting), homogene izodizomije (kromosomskog imprintinga) i oštećenja ekspresije gena u centru za otiskivanje .
Genetsko otiskivanjezasnovan na mehanizmu specifične metilacije citozinskih baza molekule DNK, koja isključuje transkripciju gena (vidi Poglavlje 28).
Trenutno se pretpostavlja da je oko 200 gena podložno takvom imprintingu i da imaju tkivno specifičnu monoaleličku ekspresiju, a pronađena su tri klastera gena lociranih u kritičnim regionima hromozoma (7q32, 11p15 i 15q11.2-q13) i koji imaju direktan uticaj na razvoj nasljedne patologije. takav mehanizam, uključujući tumore.
Genomski imprintingna osnovu homogene izodizomije na specifičnom hromozomu majčinskog ili paternalnog porekla. Uočava se u nizu nasljednih i kongenitalnih bolesti, kao što su: Angelmanovi sindromi (uzrokovani homogenom izodizomijom na 15 roditeljskom kromosomu) i Prader-Willi (isodizomija hromozoma majke 15); prolazna neonatalna dijabetes (kromosom 6); Srebrni Russellov sindrom (majčinski hromozom 7); sindrom
Beckwith-Wiedemann (očinski hromozom 11); odloženi fizički i motorički razvoj, hipotenzija i prijevremeni seksualni razvoj (majčinski hromozom 14); intrauterini zastoj rasta (IUGR) fetusa (majčinski hromozom 7), kao i IUGR sa ograničenim placentnim mozaicizmom (majčinski hromozom 16).
Efekat genomskog imprintinga se javlja i tokom mehurića. Postoje 4 vrste prekršaja:
Androgeneza ili dvostruki set hromozoma (2n): dve sperme sa X hromozomima i jaje bez jezgra;
Genogeneza ili dvostruki set hromozoma (2n): jajne ćelije sa duplim setom hromozoma, spermatozoidi nisu uključeni u oplodnju;
Android ili trostruki set hromozoma (3n): 2 očinske i 1 majčinska;
Ginoidni ili trostruki set hromozoma (3n): 2 majčinska i 1 očinska.
Utiskivanje kao rezultat grešaka u ekspresiji gena u centru za otisak
javlja se i kod Angelmanovih i Prader-Willijevih sindroma. To je zbog činjenice da oba roditelja prenose potomku pacijenta gene koji nose njihove specifične osobine, tj. geni oca i majke se aktiviraju ili potiskuju od strane potomaka na različite načine (tzv geni).Štaviše, u liniji majke i oca, roditeljski poremećaji ekspresije gena su nasleđeni u centru za otisak.
Trenutno je izolirano najmanje 60 utisnutih gena (ili njihovih transkripata) na 1, 5-7, 11, 13, 15, 19, i 20 autosoma i X kromosoma.
Treba napomenuti da su u Angelmanovim i Prader-Willijevim sindromima pronađeni geni blisko locirani, ali suprotno utisnuti. Zvali su ih geni kandidata za bolesti(u porodicama sa ponovljenim slučajevima ovih sindroma). Štaviše, kandidati genoma Angelmanovog sindroma izraženi su isključivo na majčinom hromozomu, ali su potisnuti na očinski hromozom, dok su kandidati geni Prader-Willi sindroma izraženi na očinskom hromozomu, ali su potisnuti na majčinski hromozom (vidi Poglavlje 28).
Dakle, otisak je rezultat kvalitativnih, a ne kvantitativnih promjena u nasljednom materijalu.
Krajem XX veka. takođe je identifikovao ranije nepoznatu klasu bolesti sa netradicionalnim nasledstvom - ekspanzivne bolesti
tidny repetitionspovezan sa povećanjem broja kodirajućih i nekodirajućih nukleotidnih sekvenci kao rezultat dinamičkih mutacija. Dinamička mutacija se „kreće“ iz stanja fenotipski ne manifestovane premutacije u stanje fenotipski manifestovane potpune mutacije. Takve mutacije su u osnovi brojnih teških nasljednih neurodegenerativnih bolesti (27 nosologija; vidi poglavlje 27). Na primer, ponavljanje CGT je tipično za Martin-Bell sindrom (Xq23), ponavljanje GHZ za drugu varijantu ovog sindroma (Xq28); CTG ponoviti - za spin-bulbar mišićnu atrofiju (Xq11-12); CAG - za miotoničnu distrofiju (19q13.3); CTG za Huntingtonovu koreu (4p16.3); CTG - za spinocerebralnu ataksiju, tip I (6p21.3); CTG - za Machado-Joseph-ovu bolest (14q32.1); TTC - za Friedreichovu ataksiju (9p13). U svim slučajevima postoji postupna akumulacija (ekspanzija) kritičnog broja ponavljanja nukleotida.
Paralelno sa širenjem u svakoj narednoj generaciji povećava se ozbiljnost bolesti - anticipacija.
INDIKATORI INTERAKCIJA GENA KAO DODATNI POJMOVI GENOMIKE I PROTOMIKE
Prema zakonima genetike, roditeljski i materinski genomi, ujedinjeni u genotipu jedne ćelije i celog organizma, međusobno komuniciraju u ontogenezi, stvarajući individualnu spoljašnju i unutrašnju karakteristiku, fenotip organizma.
Postoje tri grupe mehanizama interakcije između roditeljskih gena i, prema tome, tri grupe indikatora vezanih za dodatne koncepte genomike i proteomike. To su interakcije između:
Alelni geni; karakteriziraju ih: dominacija, recesija, domi- nalnost, nepotpuna i uslovna dominacija, nadmoćnost;
Nonallelic geni; karakterišu ih: epistaza, komplementarnost, polimeri;
Odvojeni genom i genotip (kao sistem gena); karakterišu ih: izražajnost, penetracija, efekat položaja,
genokopija, efekt pleiotropije, alelna serija i fenokopija.
Razmotrite ove mehanizme kao dodatne koncepte genomike i proteomike.
Mehanizmi interakcije između alelnih gena
Dominacija i udubljenje
Ako funkcionalno stanje jednog alela (na primjer, majčinog alela) ne zavisi od stanja drugog alela (alel paternal), potomak će pokazati osobinu koju kontroliše majčinski alel. Takav gen i njegova osobina se nazivaju dominatnymi.
Prvi opis efekta dominacije odnosi se na 1905. godinu, kada je porodica pacijenta sa brahidaktilijom označena kratkom pesnicom. Drugi primjeri su: bijeli uvojak, " night blindness", Habsburgova usna, polydactyly (multipawedness), syndactyly (fuzija mekog ili koštanog tkiva falanga), arachnodactyly (" paukove prste "), hondrodistrofija, kao i brojni oblici autosomno dominantnih bolesti, na primjer, Huntingtonova koreja (4p16.3).
Ako funkcionalno stanje jednog alela (na primjer, majčinog alela) zavisi od stanja drugog alela (paternal alela), potomak će ispoljavati osobinu koju kontroliraju i majčinski i očinski aleli istovremeno. Istovremeno se nazivaju i sam gen i osobina koju kontroliše recesivan.
Primjeri nasljeđivanja recesivnih osobina i fenotipa: albinizam, meka, ravna kosa, prćast nos, svijetle oči, rezus-negativna krvna grupa I, nesposobnost da se osjeti ukus feniltiokarbamida; brojni oblici autosomno recesivnih bolesti, kao što su fenoketonurija genokopija (12q24.2; 4p15.1).
U nekim slučajevima, dominacija i recesivnost gena loše koreliraju sa dominacijom i recesivnošću likova. Na primjer, mongoloidni epicant kontrolira dominantni gen, a Bushmen i Hottentots recesivni gen.
Drugi primjer je gore spomenuti gen i znak ćelavosti, koji se manifestira kod muškaraca kao dominantnih, a kod žena kao recesivnih. Kako se ispostavilo, takav mehanizam je posljedica djelovanja hormona, tj. postoji nasleđe koje zavisi od spola (pod kontrolom pola).
Nepotpuna dominacija
O nepotpunoj dominaciji,ili međudjelovanje gena (manifestacija simptoma), kažu oni kada slabe djelovanje dominantnog gena u prisustvu recesivnog gena, tj. u heterozigotama. Međutim, ne može se izvući jasna granica između interakcije i dominacije s jedne strane, kao i međudjelovanja i recesije s druge strane. Na primer, pigmentacija ljudske kože varira od bele do albino do crne u crnom. Od brakova između belaca i crnaca rođeni su mulati, koji imaju srednju boju kože.
Drugi primjer nepotpune dominacije su razlike u 6 tipova pjevajućih glasova, koje kontrolira jedan alelni par. Konkretno, bariton i mezo-sopran su uočeni samo u heterozigotima, dok su za homozigote karakteristični tenor i bas, alt i sopran.
U daljim istraživanjima, međutim, pronađena je: pigmentacija kože i tip glasa pjevanja određeni su ne samo ovim mehanizmom interakcije, već i faktorima koji su međusobno neovisni: utjecaj spolnih hormona, efekt polimernog, seksualnog ili seksualnog nasljeđivanja.
Uslovna (nestabilna) dominacija
Kada je nestabilna ili uvjetna dominacijamanifestacija svojstva u heterozigotama zavisi od genotipa i spoljašnjih uslova (modifikujući efekat genotipa na glavni gen, penetracija gena, položaj gena u hromozomu, uticaj temperature).
Co-dominance
Ako su alelni geni jednako aktivni (posjeduju istu dominantnu aktivnost), onda ovo codominance.Njegov klasičan primer je nasleđivanje IV krvne grupe (prema ABO sistemu), koje su određena sa tri alela koji se nalaze u 9. hromozomu (pluralitet alela). Među njima su dva dominantna alela (I A i! B) i jedan recesivni alel (I 0). Kombinacija ovih parova u paru daje 4 krvne grupe:
- prva grupa- prisustvo dva identična recesivna alela - I 0 I 0 (homozigot), određivanje prisustva alfa i beta antitela u krvnom serumu;
- druga grupa- prisustvo dva identična dominantna alela I A I A (homozigot) ili dva različita alela I A I 0 (heterozigot), uzrokujući prisustvo beta-antitela u serumu;
- treća grupa- prisustvo dva identična dominantna alela M B (homozigota) ili dva različita alela M0 (heterozigota); alfa antitela su prisutna u krvnom serumu;
- četvrta grupa- prisustvo dva različita dominantna alela! Oh! B (heterozigota); nema antitijela u serumu, oba alela stupaju u interakciju s istom silom, neutralizirajući jedni druge.
Još jedan primjer kodomancije je nasljeđivanje anemije srpastih stanica, koja je autosomno recesivni poremećaj (11p15). U ovom slučaju, uočava se homozigotnost (2 patološka alela jednog gena koji kontrolira sintezu defektnog hemoglobina). Takvi homozigoti imaju karakteristične simptome, ali su imuni na malariju, jer malarijski plazmodij nije reprodukovan na defektnom hemoglobinu.
Međutim, u heterozigotnom organizmu, normalni i neispravni aleli istog gena su istovremeno prisutni. Štaviše, oba alela daju isti dominantni efekat, i stoga su dva tipa hemoglobina (normalna i abnormalna) istovremeno sintetizovana u ćelijama. Takvi heterozigotni nosioci patološkog gena nemaju simptome anemije srpastih ćelija, ili se manifestiraju u blagom obliku i samo u uvjetima nedostatka kisika.
Overdominance
U nekim slučajevima, aleli koji su u heterozigotnom stanju, fenotipski su jači od alela koji su u homozigotnom stanju. (efekt overdominance).Takva manifestacija je slična efekat heterozisau biljkama (hibridna snaga ili snaga). Dakle, u slučaju brakova između predstavnika različitih rasa, zdravstveni pokazatelji njihovih potomaka su superiorniji od onih samih roditelja: djeca imaju višu vitalnost, duži životni vijek, itd.
Mehanizmi interakcije između nealelnih gena
Epistasis
Epistasis- potiskivanje djelovanja gena lociranog u jednom nealelnom paru, djelovanjem gena iz drugog nealelnog para, na primjer, supresija gena A gena B, tj. A\u003e B ili A\u003e bb. Razlikovati dominantnu i recesivnu epistazu.
Dominantna epistaza:dominantni alel jednog nonallelnog para, koji je u homozigotnom (AA) ili heterozigotnom (Aa) stanju, potiskuje ispoljavanje nelealelnog na njegov dominantni alel drugog alelnog para, koji je u AA ili Aa stanju. Nazivaju se dominantni geni epistatski geniili supresori (inhibitori). Oni mogu biti i dominantni i recesivni. Nazivaju se potisnuti geni hipostatski geni.
Ako geni pronađeni u drugim nealelnim parovima pojačavaju dominantni efekat epistatičkih gena, onda se oni nazivaju geni modifikatora(pojačavači).
Ovaj tip interakcije je karakterističan za nealelne gene koji su uključeni u regulaciju ontogeneze, na primjer, geni za imuni odgovor (mreža gena - 2190 gena; vidi poglavlje 15) ili geni eritropoeze (mreža gena - 200 gena).
Postoje dvije opcije za dominantnu epistazu:
Homozigoti sa recesivnim alelima (aa) razlikuju se u fenotipu od homozigota sa dominantnim alelima (AA);
Homozigoti za dominantne alele (AA) se ne razlikuju u fenotipu od homozigota za recesivne alele (aa).
Recesivna epistazamanifestuje se u činjenici da recesivni alel jednog gena suprimira djelovanje ne-alelnog dominantnog gena (aa\u003e B), a komplementarnost je uočena između dominantnih gena (vidi dolje). Primjer recesivne epistaze kod ljudi je "Bombajski fenomen" povezan s rađanjem djece s I (I 0 I 0) i IV (I A I B) krvnim skupinama od roditelja s I (I 0 I 0) i II (IAI °) skupinama. krvi, dok teoretski od takvih roditelja treba roditi djecu sa I (I 0 I 0) ili II (IAI 0) krvnim grupama. Fenomen se može objasniti ili prisutnošću rijetke heterozigotne varijante treće krvne grupe (IBI 0) koju nije prepoznao jedan od roditelja, ili prisutnošću u genotipu djeteta s recesivnim modifikatorskim genima IV (I i I B) krvne grupe koji suzbijaju ekspresiju antigena u homozigotnom stanju. eritrociti koji se nalaze na površini, tj. daju nepredvidivi fenotipski efekat.
Pored recesivne epistaze, istaknuta je dvostruka recesivna epistaza;tokom recesivnih gena ima svoju fenotipsku manifestaciju, au dvostrukim homozigotima recesivni aleli međusobno potiskuju: aa\u003e bb, bb\u003e aa.
Komplementarnost
Komplementarnost- tip interakcije najmanje dva dominantna nealelička gena iz nekoliko parova sa različite kombinacije dominantni i recesivni aleli, doprinoseći razvoju novih osobina osim roditeljskih varijanti.
Postoje tri tipa komplementarnosti:
Dominantni aleli (AB) se razlikuju po fenotipskoj manifestaciji;
Dominantni aleli (AB) su slični u fenotipskoj manifestaciji;
U dominantnim (A) i recesivnim (a) alelima nekoliko nealekularnih parova postoji nezavisna fenotipska manifestacija.
Na primjer, kod osobe, normalni sluh je uzrokovan interakcijom nekoliko parova nealelnih gena, ali najmanje jedan dominantni alel mora biti prisutan u parovima. Ako se ispostavi da je osoba recesivni homozigotni (najmanje jedan par nealelnih gena), onda će on biti gluv.
Drugi primjeri komplementarnosti su fenotipovi pacijenata oboljelih od raka s retinoblastom i nefroblastomom (vidi poglavlje 25).
Polimerna i kvantitativna karakterizacija
Polimer- to je uslovljenost osobine ili fenotipa interakcijom gena lokalizovanih u nekoliko nealekularnih parova i koji daju isti efekat. Ovi geni se zovu polimerni geniili poligeni. Stepen ispoljavanja svojstva (fenotip) zavisi kako od broja dominantnih gena u nealelnim parovima, tako i od broja nealelnih parova.
Takvi znakovi se zovu kvantitativne osobine.Oni se značajno razlikuju od kvalitativnih karakteristika.
Ako su kvantitativna svojstva kontrolisana genima nasleđenim poligenski, a pojavljuju se u mnogim državama kao prelazni oblici, onda kvalitativni znakovikontrolišu genski nasleđeni monogeni i pojavljuju se samo u alternativnim stanjima bez prelaznih oblika.
Primeri kvantitativnih osobina su: gore pomenuta varijabilna pigmentacija kože kod ljudi i prisustvo srednje boje kože u mulatama.
Drugi primjeri kvantitativnih osobina su: nivo (stanje) zdravlja osobe, očekivano trajanje života, intelektualne sposobnosti, težina i dužina
Posljednjih godina je istaknut fenomen interakcije između brojnih nealelnih gena: kumulativni polimeri.U ovom slučaju govorimo o aditivnom (sumirajućem) djelovanju gena, od kojih svaki ima (često mali) učinak na osobinu. To je kumulativni polimer koji formira gore pomenute genske mreže koje kontroliraju značajan dio kvantitativnih osobina organizma.
Među genima koji utiču kvantitativni atribut, postoji jedan glavni gen i niz gena koji su slabiji u odnosu na njega (poligeni). Djelovanje glavnog gena je ponekad značajno veće nego kod drugih gena, a osobina koju kontrolira glavni gen nasljeđuje se kao Mendelov (monogensko nasljeđivanje), a osobine kontrolisane poligenima nasljeđuju se prema poligenskoj varijanti. Primjer je nasljeđivanje patuljastog uzroka glavnog gena u slučaju akondroplazije, dok je kod normalne populacije ljudski rast određen aditivnim učinkom poligena.
MEHANIZMI INTERAKCIJE IZMEĐU
ODVOJENI GEN I GENOTIP
Izražajnost i penetracija
Ove koncepte je prvi put uveo 1926. godine N.V. TimofeevRessovskim i 0. Vogt opisuju različite manifestacije znakova i gena koji ih kontrolišu. Izražajnostpostoji stepen manifestacije (varijacije) iste osobine kod različitih osoba sa genom koji kontroliše ovu osobinu. Postoji niska i visoka izražajnost. Razmotrimo, na primer, različitu težinu rinitisa (curenja nosa) kod tri različita pacijenta (A, B i C) sa istom dijagnozom 0RVI. Kod pacijenta A, rinitis je izražen u blagom stepenu ("njuškanje"), što omogućava da se rukom drži jedna maramica tokom dana; pacijent ima B rinitis izražen u umjerenim (dnevno 2-3 maramice); Pacijent C ima visok stepen rinitisa (5-6 maramica). Kada govorimo o ekspresivnosti nije samo jedna osobina, već bolesti općenito, liječnici često ocjenjuju stanje pacijenta kao zadovoljavajuće ili umjereno, ili ozbiljno,
tj. u ovom slučaju, koncept ekspresivnosti je analogan konceptu "ozbiljnosti bolesti".
Penetrance- to je vjerovatnoća ispoljavanja iste osobine kod različitih pojedinaca s genom koji kontrolira tu osobinu. Penetracija se mjeri u postotku pojedinaca sa specifičnom osobinom ukupnog broja osoba koje nose gen koji kontrolira tu osobinu. 0na je nepotpuna ili potpuna.
Primer bolesti sa nepotpunom penetracijom je i dalje isti rinitis sa 0RVI. Dakle, možemo pretpostaviti da pacijent A nema rinitis (ali postoje i drugi znaci bolesti), dok pacijenti B i C imaju rinitis. Stoga je u ovom slučaju penetracija rinitisa 66,6%.
Primjer bolesti s punom penetracijom - autosomno dominantan chorea Huntington(4p16). Najčešće se javlja kod osoba u dobi od 31 do 55 godina (77% slučajeva), kod ostalih pacijenata u različitim godinama: iu prvim godinama života, iu 65, 75 i više godina. Važno je naglasiti da ako se gen ove bolesti prenosi na potomka jednog od roditelja, bolest će se nužno manifestirati, što je potpuna penetracija. Istina, pacijent ne živi uvijek do ispoljavanja Huntingtonove koreje, umirući od drugog uzroka.
Učinak položaja
Drugi tip zavisnosti gena od genotipa - efekt pozicije.Otvorio ga je A. Stertevant (1925). Suština efekta je u promeni ekspresije gena sa promenom njegovog položaja (pozicije) u hromozomu (u nizu nukleotidnih sekvenci).
Prema savremenim konceptima, efekat pozicije nije povezan sa kršenjem strukture gena: on i njegova promotorska regija su sačuvani kao jedinica transkripcije. Shodno tome, efekat pozicije je epigenomski događaj, određen trima uslovima:
U promotoru je započeta transkripcija;
Regulatorni elementi (pojačivači i prigušivači) koji sadrže mesta za vezivanje transkripcionog faktora povećavaju specifičnost transkripcionog kompleksa na promotoru;
Organizacija kromatina u području lokusa doprinosi povećanoj osjetljivosti na djelovanje nukleaza.
Nivo ekspresije zavisi od lokacije gena u genomu: bilo u područjima kondenzovanog heterohromatina, bilo u područjima
undecondensed chromatin (euchromatin), koji se dekondenzira u interfazi, sadrži većinu gena i replicira se na početku S-faze. S druge strane, heterohromatin se kondenzira kroz cijeli stanični ciklus, replicira se na kraju S faze i sadrži uglavnom ponavljajuće sekvence. Centromerni regioni hromozoma sastoje se od strukturnog heterohromatina ili gusto kondenzovane hromatina koji sadrži ponavljajuće sekvence.
Zbog ovih posebnosti u formiranju strukturalnih preraspodjela kromosoma, njihovi prekidi dovode do promjene položaja gena, što je praćeno promjenom u njihovoj ekspresiji (Sl. 22). Dominantni gen A, koji se pokazao blizu mjesta rupture kromosoma, u većoj mjeri gubi svoj utjecaj od gena B, koji se nalazi dalje od mjesta rupture. Drugim rečima, slabljenje efekta dominantnog gena proporcionalno je rastojanju između njega i tačke prekida hromozoma.
U drugim slučajevima, kromosomska pregradnja može:
Odvojiti transkripcionu jedinicu od regulatorne regije, potpuno neutralizirajući njen učinak na gen (odsustvo pojačivača smanjuje ili uklanja transkripciju u odgovarajućem tkivu); zauzvrat, odvajanje gena i njegovog prigušivača će dovesti do abnormalnog povećanja ekspresije;
Premjesti gen u regiju pojačivača drugog gena, koji uzrokuje neadekvatnu ekspresiju (na primjer, u Burkittovom limfomu, translokacija pomiče c-myc gen pod kontrolom pojačivača imunoglobina);
Premesti gen i njegove regulatorne sekvence u drugi gen, koji smanjuje nivo ekspresije prvog gena;
Da izazove mozaički efekat položaja (na primer, crvena i bela boja očiju u Drosophili).
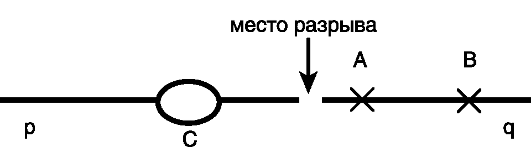 Sl. 22Učinak položaja u slučaju rupture kromosoma:
Sl. 22Učinak položaja u slučaju rupture kromosoma:
C - centromera; p i q - kratka i duga ramena kromosoma; X - gen
locus; A i B - dominantni geni
Drugi primer manifestacije efekta situacije - pojedinci sa krvnim grupama na sistem Rh. Posebno, pojedinci sa kompleksom gena CDE / cDe imaju iste gene kao i pojedinci sa cDe / CDE genskim kompleksom (gdje je veliko slovo dominantni gen, a veliko slovo recesivni gen). Međutim, prvi ima mnogo antigena E i malog antigena C, a drugi imaju suprotno, što se objašnjava nejednakom lokalizacijom gena C i E na istom hromozomu ili na različitim hromozomima.
Drugi primjer je autosomno dominantna lice-skapularna mišićna distrofija tipa 1A, uzrokovana brisanjem varijabilnog područja u subtelomernom dijelu dugog kraka kromosoma (koji se nalazi u 4q35 segmentu), koji ne sadrži gene, ali ima nukleotidne sekvence veličine 20-250 kb. Brisanje ovih sekvenci dovodi do promjene u strukturi heterohromatina u ovom području kromosoma i posredovane inhibicije transkripcije obližnjeg gena (FRG1) lociranog proksimalno od točke rupture kromosoma.
Nazovimo niz nasljednih bolesti koje su povezane sa efektom situacije. Među njima su:
Aniridijum kao rezultat delecije gena lociranog na lokusu
Holoprocephalus - mutacija u SSN genu (7q36);
Satre-Kotzen craniosynostosis - mutacija u TWIST genu (7p21);
Cephalopolisindactyly Greig kao rezultat mutacije SLIZ
Riegerov sindrom - PITX2 gen (4q25-q27);
XY preinačenje pola - mutacija u genu SRY (Xq).
Genokopiranje i njegovi uzroci
Ista osobina (grupa osobina) je uzrokovana različitim genetskim uzrocima (ili heterogenosti). Takav učinak, na prijedlog njemačkog genetičara H. Naftheima, dobiven je sredinom 40-ih godina 20. stoljeća. ime genocopying.Poznate su tri grupe uzroka genokopije.
Uzroci prve grupekombinira heterogenost zbog multikulturalnosti ili djelovanja različitih gena lociranih u različitim lokusima na različitim kromosomima. Na primjer, među nasljednim metaboličkim oboljenjima kompleksnih šećera - glukoznih aminoglikana, izdvaja se 19 tipova (podtipova) mukopolisaharidoza. Sve vrste
zahvaćeni su defekti različitih enzima, ali se manifestiraju istim (ili sličnim) simptomima gargoylic dysmorphismili fenološki tip zvona Quasimodo - protagonist romana „Notre-Dame de Paris“ klasika francuske književnosti Viktora Huga. Sličan fenotip se često uočava kod mukolipidoze (poremećaji metabolizma lipida).
Drugi primjer multilocusa je fenilketonurija. Ne samo klasični tip, zbog nedostatka fenilalanin-4-hidroksilaze (12q24.2), nego i tri atipična oblika su istaknuti: jedan je uzrokovan nedostatkom dihidropteridinereduktaze (4p15.1), a još dva - nedostatkom piruvoil tetrahidropterin sintetaze i tetrahidrobiopterin deficijencije geni koji još nisu identifikovani).
Dodatni primjeri multilocusnosti: glikogenoza (10 genokopija), Ellers-Danlosov sindrom (8), Reklingausenova neurofibromatoza (6), kongenitalni hipotiroidizam (5), hemolitička anemija (5), Alzheimerova bolest (5), Barde-Biedlov sindrom (3), rak dojke (2).
Uzroci druge grupekombinuje intralokalnu heterogenost. To je ili zbog toga multipli alelizam (vidi poglavlje 2), bilo putem genetski spojeviili dvostruki heterozigoti sa dva identična patološka alela u identičnim lokusima homolognih kromosoma. Primer potonjeg je heterozigotna beta-talasemija (11p15.5), koja se formira kao rezultat delecija dva gena koji kodiraju beta lanac globina, što dovodi do povećanog sadržaja hemoglobina HbA 2 i povećanog (ili normalnog) nivoa hemoglobina HbF.
Uzroci treće grupekombinuje heterogenost zbog mutacija u različitim tačkama istog gena. Primjer je cistična fibroza (7q31-q32), koja se razvija zbog prisutnosti gotovo 1000 točkastih mutacija u genu odgovornom za bolest. Sa ukupnom dužinom gena cistične fibroze (250 hiljada N. p.) Očekuje se da će detektovati do 5.000 takvih mutacija u njemu. Ovaj gen kodira protein koji je odgovoran za transmembranski prenos jona hlora, što dovodi do povećanja viskoznosti sekrecije egzokrinih žlezda (znojenje, salivarno, sublingvalno, itd.) I blokiranje njihovih kanala.
Drugi primjer je klasična fenilketonurija, zbog prisutnosti 50 točkastih mutacija u genu koji kodira fenilalanin-4-hidroksilazu (12q24.2); Sve u svemu, ova bolest bi trebala otkriti više od 500 točkastih mutacija gena. Većina njih se pojavljuje
zbog polimorfizma duž dužine restrikcijskog fragmenta (RFLP) ili broja tandemskih ponavljanja (VNTP). Utvrđeno je da je glavna mutacija gena fenilketonurije u slavenskim populacijama R408 W /
Pleiotropski efekat
Navedena dvosmislenost prirode veza između gena i karaktera također je izražena u pleiotropski efekatili pleiotropsko djelovanje, kada jedan gen uzrokuje stvaranje više znakova.
Na primjer, autosomno recesivni ataksija telangiektazijski gen, ili louis Bar sindrom(11q23.2) je odgovoran za istovremenu destrukciju najmanje šest telesnih sistema (nervni i imunološki sistem, koža, respiratorne i gastrointestinalne sluznice i konjunktiva oka).
Drugi primjeri: gen bardet-Beadle sindrom(16q21) uzrokuje demenciju, polidaktilnost, gojaznost, degeneraciju pigmenta mrežnice; Gen Fanconi anemije (20q13.2-13.3), koji kontroliše aktivnost topoizomeraze I, uzrokuje anemiju, trombocitopeniju, leukopeniju, mikrocefaliju, aplaziju radijusa, hipoplaziju metakarpalne kosti prvog prsta, malformacije srca i bubrega, hipospadiju, age spots kože, povećana krhkost kromosoma.
Izdvojite primarnu i sekundarnu pleiotropiju. Primarna pleiotropijazbog biohemijskih mehanizama djelovanja mutantnog enzimskog proteina (na primjer, nedostatak fenilalanin-4-hidroksilaze u fenilketonuriji).
Sekundarna pleiotropijazbog komplikacija patološkog procesa koji se razvio kao rezultat primarne pleiotropije. Na primjer, zbog pojačanog stvaranja krvi i hemosideroze parenhimskih organa, pacijent sa talasemijom razvija zadebljanje kostiju lobanje i hepatolijalni sindrom.
Allelic series
Tačkaste mutacije jednog gena mogu uzrokovati razvoj ne samo jednog, već i različitih kompleksnih simptoma. U drugom slučaju, igra ulogu alelne serije istog gena.Na primer, mutacije gena adrenoreceptora povezane sa uzrokom X hromozoma kennedyjeva bolest,ako uhvate regiju trinukleotidnih ponavljanja u egzonu 1, ali mogu dovesti do toga syn
morris Dromaili feminizacija testisa (fenotipski - djevojka, genotipski - dječak, to jest, prisustvo ženskog fenotipa sa muškim kariotipom), ako su uhvaćene druge nukleotidne sekvence ovog gena (mutacija u domenu vezivanja androgena).
Drugi primjer heterogenosti alelne serije su točkaste mutacije gena receptora tirozin kinaze - RET, uzrokujući četiri nasljedne bolesti: porodični medularni karcinom štitne žlezde, Hirschsprungova bolest, višestruka endokrina neoplazijadva tipa - 2A (MEN2A) i 2B (MEN2V).
Iako je opisani fenomen otkriven relativno nedavno, već je poznato više od 100 monogenih bolesti uzrokovanih mutacijama jednog gena u serijama alela. Njihovi razlozi su:
Različite nukleotidne sekvence (različite tačke gena) koje kontroliraju funkcionalno različite proteinske domene;
Prisustvo u istom genu koji modifikuje mutantni alel (alelni polimorfizam);
Uticaj genetske sredine na manifestaciju mutantnog alela (uključujući interakciju mutantnog alela sa alelima jednog ili više modifikatorskih gena).
Fenokopiranje
Fenokopiranje- ovo je kopiranje kompleksa simptoma ne-nasljedne bolesti (nastale pod utjecajem faktora okoliša) kao simptomskog kompleksa nasljedne genetske bolesti.
Na primjer, simptomi endemske guše odgovaraju simptomima kongenitalnog hipotiroidizma, ali u prvom slučaju razlog je deficit (ili potpuna odsutnost) neorganskog joda u vodi za piće i hrani (tj. Fenokopija), au drugom - različiti genetski faktori (m. e. postoji genokopija).
Drugi primjer simptoma duplikacije gena i fenokopije je sljepoća. Poznato je da se vizija kontroliše grupom gena (mreža gena) koji međusobno komuniciraju tokom ontogeneze i osiguravaju razvoj i održavanje funkcionisanja očiju i mozga. Različiti razlozi za kršenje integriteta ovog sistema mogu izazvati isti rezultat (sljepoća).
Zbog toga, zbog genokopije, može doći do razvoja slepila zbog defekata sočiva određenih određenim genima, retinalnim defektima koji su definisani drugim genima, i defektima rožnice određenim trećim genima.
Kao rezultat fenokopija, usled egzogenog uticaja okoline, može se razviti slepilo zbog nepoštovanja standarda za osvetljenje stambenih i industrijskih objekata ili kao posledica povreda oka.