Моногибридное схрещування двох гетерозиготних. Регуляція експресії генів у прокаріотів. Зчеплення генів. Досліди і правило Моргана
При схрещуванні білих курей Андалузії з чорними півнями були отримані курчата з сірим оперенням. При схрещуванні сірих курей між собою було отримано потомство F 2. 1) Який генотип мають сірі кури? 2) Скільки класів генотипів і фенотипів формується в F 2? 3) Які кури F 2 дадуть нерозщеплюваних потомство?
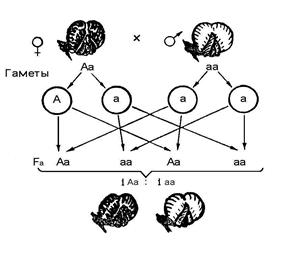
Успадкування забарвлення оперення у курей Андалузії показано на рис. 2. Хід міркувань і визначення закономірностей успадкування ознак аналогічні задачі 2. Оскільки з умов завдання випливає, що гетерозиготи мають проміжну забарвлення, генотип батьківських форм відповідає формулам: аа і АА. Схрещування білих і чорних курей дає сіре потомство F 1 c генотипом Аа. Гетерозиготні кури утворюють два типи гамет. В результаті в F 2 формується 3 класу генотипів у співвідношенні 1АА: 2Аа: 1аа. На відміну від попереднього випадку фенотип гомозиготних форм відрізняється від фенотипу гетерозиготних особин і в потомстві з'являються білі, сірі та чорні кури в співвідношенні 1: 2: 1. Отже, при нЕ повному домінуванні розщеплення за генотипом відповідає розщепленню за фенотипом. На малюнку також показано, що в F 3білі і чорні кури дають однотипних курчат, а при схрещуванні гетерозиготних курей спостерігається знову розщеплення за фенотипом у співвідношенні 1: 2: 1.
відповіді:1) сірі кури мають генотип Аа; 2) в F 2 формується 3 Кламма генотипів і 3 класу фенотипів; 3) нерозщеплюваних потомствовдадут чорні і білі кури.
На прикладі цих двох завдань можна зробити висновки:
1. При схрещуванні гомозиготних рослин (або тварин) і їх гібридів між собою спадкування ознак підпорядковується законам Менделя.
2. Фенотип гібридів F 1 вказує на напрямок і ступінь домінування генів (ознак).
3. вищепленію форм з рецесивним ознакою можливо тільки у потомства гетерозиготних форм, в той час як гомозиготи дають стабільне потомство
Мал. 11.Успадкування забарвлення оперення у курей Андалузії
за типом неповного домінування
.Завдання 3. Аналізуюче схрещування.
У анализирующем схрещуванні рослин гороху з пурпуровими квітками було отримано 18 рослин з пурпуровими і 18 рослин з білими квітками. 1) Який генотип має аналізоване рослина? 2) Скільки типів гамет утворює ця рослина? 3) Яка частина рослин, отриманих в анализирующем схрещуванні, дасть нерозщеплюваних потомство?
У цьому завданню необхідно визначити генотип батьківських форм завдяки знанню особливостей аналізує схрещування. Оскільки за визначенням аналізує схрещування - це гібридизація з гомозиготною рецесивною формою, то генотип батьківської форми - аа, Що відповідає белоцветковая фенотипу. Для визначення генотипу материнської пурпурноцветковой форми необхідно згадати, які результати дає аналізує схрещування з домінантною гомозиготною і гетерозиготною формою:
Р ♀ АА х ♂ аа Р ♀ Аах ♂ аа
F 1 Аа F 1 Аа аа
Як видно зі схеми, якщо рослина з домінантним фенотипом гомозиготное (АА), все гібриди мають домінантний ознака; а при схрещуванні з гетерозиготною формою в потомстві спостерігається розщеплення в співвідношенні 1: 1.
За умовами завдання в результаті аналізує схрещування в потомстві спостерігалося 18 пурпурноцветкових і 18 белоцветкових форм, що відповідає розщепленню 1: 1 і вказує на гетерозиготність материнської форми. Далі хід завдання проілюстровано на рис. 3. Анализируемая форма утворює два типи гамет. При злитті гамет утворюються гетерозиготні пурпурноцветковие і гомозиготні білоквіткові рослини. Як було показано в попередніх прикладах, стабільне потомство дають тільки гомозиготні рослини, а гетерозиготні демонструють розщеплення в наступних поколіннях. Отже, нерозщеплюваних потомство дадуть тільки 18 рослин з білими квітками.
Мал. 12. Ще Одне схрещування гібрида F 1 з рецесивною
батьківською формою (аналізує схрещування)
відповіді: 1) аналізоване рослина має генотип Аа; 2) аналізоване рослина утворює два типи гамет; 3) в результаті аналізує схрещування 1/2 потомства, тобто 18 рослин, будуть гомозиготними і дадуть нерозщеплюваних потомство.
завдання 4. Дигибридное схрещування.При схрещуванні сорту томатів, що мають червоні круглі плоди з рослинами, що мають жовті грушоподібні плоди, були отримані красноплодной гібриди з круглими плодами. В результаті самозапилення було отримано 80 гібридів F 2. 1) Яка частина рослин F 1 була дігетерозіготной? 2) Скільки типів гамет утворюють рослини F 1? 3) Скільки класів генотипів утворюють гібриди F 2? 4) Яке розщеплення за фенотипом спостерігається в F 2? 5) Скільки рослин F 2 матиме червоні грушоподібні плоди?
Для вирішення цього завдання необхідно визначити генотипи батьківських форм. Позначимо гени, що визначають ознака кольору плодів буквою А, А форми плодів - буквою В. Напрямок та ступінь домінування визначимо за фенотипом гібридів F 1. Оскільки у гібридів першого покоління виявилися ознаки червоного забарвлення і круглої форми, ці ознаки і відповідні їм гени проявляють повне домінування. Позначимо гени томатів наступним чином:
А - червоне забарвлення плодів, а - жовте забарвлення плодів;
В - кругла форма, в - грушоподібна форма.
Схрещувати рослини диплоїдні, тому визначимо генотип материнської форми з червоними круглими плодами як ААВВ, А батьківській з жовтими грушоподібними плодами - аавв. У розглянутому прикладі ознаки успадковуються незалежно і розподіл генів пов'язано з незалежним розходженням двох пар гомологічних хромосом в профазі мейозу. Для кращого розуміння матеріалу рекомендується алельних гени, локалізовані в різних парах хромосом, підкреслювати різними лініями, що символізують приналежність до різних гомологічним парам (рис. 5). При утворенні гамет в мейозі в кожну гамет випадковим чином розподіляється по одній хромосомі з кожної пари гомологічних хромосом і по одному аллельному гену відповідно. гомозиготні батьківські форми утворюють по одному типу гамет. При їх злитті утворюються рослини, гетерозиготні по двох парах генів, - дигетерозиготи. рослини F 1 однакові по кожній парі ознак, тобто проявляється
I закон Менделя.
Дігетерозіготние рослини F 1 утворюють 2 + 2 = 4 типи гамет, при цьому поєднання хромосом і генів різних пар в гаметах випадкове і равновероятное. Для визначення генотипів і фенотипів гібридів F 2 необхідно побудувати грати Пеннета. Решітка складається з 16 (4 2) осередків, поява відповідного їм генотипу равновероятно. Підрахунок показує, що при дигибридном схрещуванні утворюється 9 (3 2) генотипів. За фенотипом потомство розподіляється на 4 класу в співвідношенні:
Рослини з червоними грушоподібними плодами мають генотип А-ст і становлять 3/16 потомства, тобто 15 рослин.
відповіді: 1) все рослини F 1 дігетерозіготни; 2) гібриди першого покоління утворюють 4 типи гамет; 3) в F 2 утворюється 9 генотипів і 4 класу фенотипів в співвідношенні 9: 3: 3: 1; 5) в F 2 з'являться 15 рослин з червоними грушоподібними плодами.

Мал. 5. Успадкування ознак кольору і форми плодів у томатів
Досліди Мендель проводив на горосі. При схрещуванні сортів гороху, що мають жовті й зелені насіння (схрещувалися гомозиготні організми або чисті лінії), все потомство (тобто гібриди першого покоління) виявилося з жовтими насінням. Протилежний ознака (зелене насіння) як би зникає. Виявлена закономірність отримала назву правило однаковості (домінування) гібридів першого покоління (абоперший закон Г. Менделя ).
Досліди по схрещуванню записують у вигляді схем:
А - ген жовтого забарвлення
а - ген зеленого забарвлення
Р - (parents - батьки)
F - (filii - діти)
Р ♀АА х ♂аа
F 1 Аа - 100% жовті
Отже, все гібриди першого покоління виявляються однорідними: гетерозиготними за генотипом і домінантними за фенотипом.
Таким чином, перше правило (закон) Менделя одноманітності гібридів першого покоління можна сформулювати наступним чином: при схрещуванні гомозиготних особин, які відрізняються один від одного по одній парі альтернативних ознак, Все потомство у першому поколінні одноманітно як за фенотипом, так і за генотипом
Правило розщеплення. Другий закон Менделя
Якщо схрестити гібриди першого покоління між собою, у другому поколінні з'являються особини, як з домінантними, так і з рецесивними ознаками, тобто виникає розщеплення в певному чисельному співвідношенні. У дослідах з горохом жовтих насіння виявляється в три рази більше, ніж зелених. Ця закономірність отримала назву другого закону (правило) Менделя, або закону (правило) розщеплення.
Р ♀ Аа х ♂ Аа
G (А) (а) (А) (а)
F 2 АА; Аа, Аа; аа
жовті зелені
Розщеплення за фенотипом 3: 1, за генотипом 1АА: 2Аа: 1аа
Другий закон (правило) Менделя : При схрещуванні двох гетерозиготних особин, аналізованих по одній альтернативної парі ознак (тобто гібридів), в потомстві очікується розщеплення за фенотипом 3: 1 і за генотипом 1: 2: 1.
Ді-і полігібридне схрещування. Третій закон Менделя
При дигибридном схрещуванні батьківські організми аналізуються по двох парах альтернативних ознак. Мендель вивчав такі ознаки як забарвлення насіння і їх форму. При схрещуванні гороху з жовтими і гладенькими насінням з горохом, мають зелені і зморшкуваті насіння, в першому поколінні все потомство виявилося однорідним, проявилися тільки домінантні ознаки – жовтий колір і гладка форма. Отже, як і при моногібрідномсхрещуванні тут мало місце правило одноманітності гібридів першого покоління або правило домінування.
А - ген жовтого кольору
а - ген зеленого кольору
В - ген гладкої форми
в - ген зморшкуватої форми
Р ♀ААВВ х ♂аавв
ж. гл. з. морщиться.
G (АВ) (ав)
F 1 АаВв - жовті гладкі
При схрещуванні гібридів першого покоління між собою відбулося розщеплення за фенотипом:
Р ♀ АаВв х ♂АаВв
9 частин - жовтих гладких
3 частини - жовтих зморшкуватих
3 частини - зелених гладких
1 частина - зелених зморшкуватих
З цього схрещування видно, що в другому поколінні були не тільки особини з поєднанням ознак батьків, але і особини з новими комбінаціями ознак.
Крім того, Мендель виявив, що кожна пара ознак (колір і форма) дала розщеплення приблизно в відношенні 3: 1, тобто як при моногібрідномсхрещуванні. Звідси був зроблений висновок, що кожна пара альтернативних ознак при ди- та полигибридном схрещуванні успадковується незалежно один від одного.
Третє правило або третій закон Менделя формулюється так: при схрещуванні гомозиготних особин відрізняються двома (або більше) парами альтернативних ознак, у другому поколінні спостерігається незалежне успадкування і комбінування ознак, якщо гени, що визначають їх, розташовані в різних гомологічних хромосомах.
Крім законів, Мендель сформулював дві гіпотези: факторальних і гіпотезу «чистоти гамет», за допомогою яких він спробував пояснити встановлені закономірності.
факторальних гіпотеза вказує на те, що в клітинах міститься фактор (ген), який і несе ознака. Батьки передають нащадкам не ознаки, а ці фактори.
Гіпотеза «чистоти гамет» : Організм за кожною ознакою несе два спадкових фактора (один від батька, другий від матері). Ці спадкові чинники, перебуваючи в клітинах, не зливаються один з одним і при формуванні гамет розходяться в різні гамети.
аналізує схрещування
Рецесивний аллель проявляється тільки в гомозиготному стані. Тому про генотип організму виявляє рецесивний ознака можна судити за фенотипом.
Гомозиготна і гетерозиготна особини, що проявляють домінантні ознаки за фенотипом не відрізняються. Для визначення генотипу виробляють аналізує схрещування і дізнаються генотип батьків по потомству.
Аналізує схрещування полягає в тому, що особина, генотип якої не ясний, але повинен бути з'ясований схрещується з рецесивною формою. Якщо від такого схрещування все потомство виявиться однорідним, значить аналізована особина гомозиготна, якщо ж станеться розщеплення, то вона гетерозиготна
Р ♀ АА х ♂ аа
Р ♀ Аа х ♂ аа
G (А) (а) (а)
Як видно зі схеми, при анализирующем схрещуванні для потомства гетерозиготною особини характерно розщеплення 1: 1.
Хромосомна теорія спадковості
план
1.Пол як спадковий ознака.
2. Визначення статі.
3.Наследованіе ознак, зчеплених зі статтю і обмежених підлогою.
4.Сцепленіе генів. Досліди і правило Моргана.
5.Основние положення хромосомної теорії спадковості.
Хромосомна теорія спадковості - розділ генетики про локалізацію успадкованих факторів в хромосомах клітин.
Суть її полягає в тому, що всі ознаки і властивості живих організмів визначаються генами, розташованими в хромосомах клітини в лінійному порядку. Засновником її з'явився американський генетик Томас Морган, теорія розроблена його співробітниками на початку ХХ століття.
Пол як спадковий ознака
Однією з ознак у багатьох живих організмів є підлогу (чоловічий і жіночий).
Пол - це сукупність морфологічних, фізіологічних, біохімічних і поведінкових ознак організму, які зумовлюють репродукцію (відтворення).
Пол - це генетично обумовлений ознака, тобто стать зародка визначається генетичними механізмами - комбінацією статевих хромосом. Статеві хромосоми визначають первинні статеві ознаки.
Під їх впливом формуються статеві залози (насінники і яєчники), що виділяють чоловічі і жіночі статеві гормони. Статеві гормони визначають розвиток вторинних статевих ознак тієї чи іншої статі і проявляються у організмів в період статевого дозрівання (у людини в 12 - 15 років). Вторинні статеві ознаки - відмінність в розмірах і пропорціях тіла, тембр голосу, ступінь розвитку волосяного покриву і ін.
визначення статі
У більшості організмів стать визначається в момент запліднення (сінгамно) і регулюється хромосомним набором зиготи, його називають хромосомний тип визначення статі.
У людини і ссавців жіночі особини мають в каріотипі дві Х - хромосоми (ХХ), а чоловічі Х- і У-хромосоми. Жіночі особини є гомогаметний, оскільки формують лише один тип гамет, а чоловічі - гетерогаметних, тому що утворюють два типи гамет.
У птахів і метеликів гетерогаметним підлогою є жіноча стать, а гомогаметний - чоловічий. У них статеві хромосоми позначають як Z і W.
У деяких клопів, жуків самки мають склад статевих хромосом ХХ, самці ж несуть всього одну з них - (ХО).
Абсолютно особливий тип визначення статі є у перетинчастокрилих, зокрема, у бджіл. У цих комах самки розвиваються з запліднених яєць і клітини їх тіла диплоїдні, а самці розвиваються партеногенетически з незапліднених яєць і мають гаплоїдні клітини.
Хромосомна теорія наслідування статі стверджує, що гени визначають розвиток статі, локалізовані в статевих хромосомах. У людини гени, які обумовлюють розвиток жіночої статі, знаходяться в Х - хромосомі, а гени чоловічої статі - в У - хромосомі.
Основні типи хромосомного визначення статі
Інша працювати у дрозофіли. При вивченні успадкування статі у мухи - дрозофіли було встановлено, що у самки дві однакові Х - хромосоми, а у самця Х- і У-- хромосоми. У дрозофіли гени, що визначають жіноча стать, знаходяться в Х - хромосомі, а У - хромосома генетично інертна і гени, що визначають розвиток чоловічої статі, знаходяться в аутосомах. Запліднені яйцеклітини, мають диплоїдний набір хромосом і дві Х - хромосоми дають початок самкам (2А + ХХ), а мають диплоїдний набір аутосом і одну Х - хромосому дають самців (2А + ХУ).
Досліди підтвердили, що чим більше наборів аутосом, тим більше були виражені ознаки чоловічої статі. З цього був зроблений висновок, що У - хромосома у мухи - дрозофіли не має істотного значення для визначення чоловічої статі.
Крім сінгамного способу визначення статі існує епігамний спосіб диференціювання статі, який визначається впливом умов навколишнього середовища, наприклад, у морського кільчастого черв'яка Bonnelia визначається умовами розвитку яйцеклітин (прогамний спосіб, як у коловерток).
Успадкування ознак, зчеплених зі статтю і обмежених підлогою
Зчепленими з підлогою називають ознаки, розвиток яких обумовлено генами, розташованими в статевих хромосомах.
Якщо ген знаходиться в У-хромосомі, то він успадковується у людини, ссавців - від батька до сина, а у птахів - від матері до дочки.
Якщо ген розташовується в Х-хромосомі, успадкування буде більш складним. Вперше особливості успадкування генів, зчеплених з Х-хромосомою були відкриті в дослідах на дрозофілі при вивченні успадкування забарвлення очей. Червоне забарвлення очей домінує над білою.
При схрещуванні гомозиготною червоноокої самки з білоокоїсамця в F1 всі потомство червонооке.
Р ♀ Х А Х А х ♂ХªУ
G (X А) (Xª) (У)
F 1 X А Xª х X А У
G (Х А) (Xª) (X А) (У)
F 2 Х А Х А, Х А Хª, Х А У, ХªУ
кр. кр. кр. бел.
При схрещуванні гібридів першого покоління між собою в F2 відбувається розщеплення за фенотипом: 3 червоноокі особини: 1 чернь, і білооких бувають тільки самці.
При реципрокних (зворотному) схрещуванні, коли самка гомозиготна по гену білооких, схрещується з червонооким самцем, розщеплення в F1 за фенотипом 1: 1. При цьому білооких будуть тільки самці. При схрещуванні мух F1 між собою в F2 будуть мухи з обома ознаками в рівному співвідношенні 1: 1, як серед самок, так і серед самців.
Р ♀ ХªХª х ♂Х А У
G (Хª) (Х А) (У)
F 1 Х А Хª х ХªУ
G (Х А) (Хª) (Хª) (У)
F 2 Х А Хª, ХªХª, Х А У, ХªУ
кр. бел. кр. бел.
З результатів досвіду Моргана можна зробити висновок, що гени забарвлення очей розташовані тільки в статевий Х-хромосомі. Результати схрещування залежать від того, який підлогу несе домінантний ознака.
Спадкування, зчеплене з підлогою, необхідно відрізняти від спадкування, обмеженого підлогою.
Розвиток ознак (обмежених підлогою) обумовлено генами, розташованими в аутосомах (а не в статевих хромосомах), але на прояв ознак у фенотипі сильно впливає стать, тобто розвиток ознаки залежить від впливу статевих гормонів.
Наприклад, облисіння з чола, типово для чоловіків. У чоловіків під дією чоловічих статевих гормонів (тестостерону) ген веде себе як домінантний. Ще приклад, тембр голосу - баритон і бас - характерні тільки для чоловіків.
Зчеплення генів. Досліди і правило Моргана
Вивчення зчепленого з підлогою спадкування стимулювало вивчення зчеплення між генами, які у аутосомах.
Для будь-якого організму характерно видове сталість хромосом в каріотипі. Генів, що визначають ознаки, у організмів набагато більше, ніж хромосом. Наприклад, у мухи дрозофіли 8 хромосом в соматичних клітинах, а генів близько 1000. Значить, в кожній хромосомі знаходиться багато генів.
Гени, локалізовані в одній хромосомі, утворюють групи зчеплення.
Число груп зчеплення дорівнює гаплоидному числу хромосом.
Успадкування генів, локалізованих в одній хромосомі, називається зчепленим спадкуванням.
На початку ХХ століття Т. Морган і його співробітники описали явище зчеплення генів - спільну передачу груп генів з покоління в покоління.
Досліди проводилися на мухах дрозофилах з урахуванням двох пар альтернативних ознак:
Колір тіла - сірий (В) і чорний (в)
Довжина крил - нормальні (V) і короткі (v)
У мухи - дрозофіли забарвлення тіла і довжини крил знаходяться в одній парі гомологічних хромосом, тобто відносяться до однієї групи зчеплення, що і було доведено в дослідах. Запис генотипів при зчепленні видозмінюється: генотип записується BV
(Дві рисочки означають, що організм диплоїдний).
При схрещуванні мух, що мають сірий колір тіла і нормальні крила з мухами чорного кольору і короткими крилами в F1 всі мухи мали сірий колір тіла і нормальні крила.
Запис схеми схрещування:
сер.нор. чер.кор.
G ( BV) (bv)
Провели аналізує схрещування. У першому випадку схрестили гібридного самця з сірим тілом і нормальними крилами
з чорної самкою, що має короткі крила, і в результаті схрещування отримали 2 типу нащадків, схожих на батьків в співвідношенні 1: 1.
Ген сірого кольору тіла і ген нормального будови крил передаються разом, а ген чорного кольору тіла і ген коротких крил теж разом. Цей досвід демонструє повне зчеплення. Причина його полягає в тому, що гени, які обумовлюють два різних ознаки, лежать в одній хромосомі. Це видно на схемі повного зчеплення:
G ( bv) ( BV) (bv)
Інший результат виходить, якщо для аналізує схрещування брати гібридну самку з сірим тілом і нормальними крилами і рецесивного за обома ознаками самця.
В цьому випадку з'явилося чотири типи нащадків у співвідношенні:
сірих довгокрилих - 41, 5%; сірих короткокрилих - 8, 5%,
чорних довгокрилих - 8, 5%, чорних короткокрилих - 41, 5%.
У цьому випадку має місце неповне зчеплення:
Р ♀BV ♂ bv
G ( BV) (bv)
некроссов. ( bv)
(Bv) (bV)
F 1 ═ ═ ═ ═
41,5% 41,5% 8,5% 8,5%
Причиною неповного зчеплення генів є кроссинговер (перехрещення хромосом) і обмін ділянками між гомологічними хромосомами в профазі I поділу мейозу при дозріванні статевих клітин. Кроссинговер відбувається у самок, у самців дрозофіл кроссинговер не відбувається (встановлено, що для особин гетерогаметного статі характерна більш низька частота кроссинговер).
При вивченні результатів схрещування виходить 4 фенотипу серед нащадків:
17% (8, 5 + 8, 5) особин утворилися з кроссоверних гамет
83% (41, 4 + 41, 5) особин утворилися з некроссоверних гамет.
Частота кросинговеру залежить від відстані між генами в хромосомі. Відстань між генами виражається у відсотках кросинговеру між ними і позначається в морганідах.
Вивчення Морганом зчеплення генів являє собою закономірне біологічне явище.
Гени, локалізовані в одній хромосомі, успадковуються, зчеплене, причому сила зчеплення залежить від відстані між генами. Ця закономірність отримала назву правило Моргана.
Основні положення хромосомної теорії спадковості
Основні положення хромосомної теорії спадковості зводяться до наступного:
Носіями спадкової інформації є хромо-
соми і розташовані в них гени,
Гени розташовані в хромосомі в лінійному порядку один за
іншому в певних локусах. Алельних гени займають
однакові локуси гомологічних хромосом,
Гени, розташовані в одній хромосомі, утворюють групи
зчеплення і успадковуються переважно разом. число
груп зчеплення дорівнює гаплоидному набору хромосом,
Між гомологічними хромосомами можливий обмін участ-
ками - кросинговер, який порушує зчеплення генів.
Сила зчеплення залежить від відстані між генами,
Відсоток кросинговеру пропорційний відстані між
генами. За одиницю відстані приймається 1 морганіда,
яка дорівнює 1% кросинговеру,
При неповному зчепленні в сумі ймовірність некроссовер-
них гамет (гібридів) завжди більше, ніж 50%,
При відстані в 50 морганід і більше ознаки наследуют-
ся незалежно, незважаючи на те, що локалізовані в 1 хромо-
молекулярна генетика
план
1. Етапи розвитку молекулярної генетики.
2. Генетичний код і його властивості.
3. Функціонально-генетична класифікація генів.
4. Функціональна активність генів (експресія генів).
5. Регуляція експресії генів у про - і еукаріот.
Молекулярна генетика займається вивченням структурно-функціональної організації генетичного апарату клітин і механізму реалізації спадкової інформації.
Етапи розвитку молекулярної генетики
Молекулярна генетика виділилася з біохімії та сформувалася як самостійна наука в 50-х роках минулого століття. Народження цієї науки пов'язане з рядом важливих біологічних відкриттів:
1). У 20-40 рр. ХХ століття було встановлено, що носієм спадкової інформації в клітині є молекула ДНК, а не білок, як вважали раніше. Були отримані прямі докази ролі ДНК у спадковості. Це явища трансформації, трансдукції, статевий процес у бактерій, будова вірусу, а також виявлення майже повну ідентичність хімічного складу ДНК (але не білків) у всіх представників даного біологічного виду.
Дані про видовий специфічності будови ДНК отримані на початку 50-х років Ервін Чаргафф і його співробітниками. Будь-яка ДНК складається з чотирьох типів нуклеотидів, що містять одне з чотирьох азотистих основ: аденін, тимін, гуанін, цитозин. Чаргафф з співробітниками виявив, що співвідношення між нуклеотидами, що входять до складу ДНК однакові в межах виду і розрізняються у різних видів. Крім того, число аденілових нуклеотидів дорівнює числу тіміділових, а число гуанілова дорівнює числу цітозілових. Це відкриття послужило головним ключем до виявлення структури молекули ДНК.
трансформація у бактерій - це передача від однієї бактеріальної клітини іншої спадкового матеріалу. У 1928 році Фред Гріффіт вивчав трансформацію у двох штамів пневмокока: вирулентного (що викликає загибель мишей) і авірулентние (що не викликає загибелі).
Якщо вбити хвороботворні бактерії і вводити їх разом з живими авірулентнимі, то частина тварин все одно загине. Цей досвід свідчив про те, що частина генетичного матеріалу з вірулентних бактерій проникла в авірулентние і зробила їх вірулентними. Після того як хвороботворні клітини розділили на окремі компоненти: вуглеводи, ліпіди, білки і ДНК, було виявлено, що саме ДНК наділило живі бактерії генетичною ознакою, якого у них до цього не було.
Так, при внесенні в нерухомі бактерії вірусів, розмножуються в клітинах рухомих культур, деякі з бактерій стають рухливими.
Статевий процес у бактерій - кон'югація, коли відбувається обмін молекулами ДНК, і хоча нові бактерії не утворюються, але їх спадковий матеріал змінюється, тому що відбувається рекомбінація генетичного матеріалу.
Будова вірусу або бактеріофага: Вірус складається з молекули ДНК (або РНК), укладеної в білкову оболонку. За допомогою радіоізотопів було доведено, що саме ДНК проникає в клітину, а білкова оболонка залишається зовні. В результаті утворюються нові фагів частинки. Таким чином, ДНК є носієм генетичної інформації і дає початок новим бактеріофагів.
непрямі докази: в соматичних клітинах різних органів міститься однакова кількість ДНК, вдвічі більшу, ніж в статевих. Кількість білків варіює і не завжди більше, ніж в статевих.
2) Слід етап розвитку молекулярної генетики пов'язаний з таким важливим відкриттям, як встановлення структурної організації молекули ДНК в 1953 році. Крик і Уотсон встановили, що ДНК складається з двох спірально-закручених ланцюгів.
В даний час прийнято говорити про первинної, вторинної та третинної структурі ДНК: Первинною структурою ДНК називають лінійну полінуклеотидних ланцюг, в якій мононуклеотиди з'єднані 3`, 5`- фосфоефірную зв'язками. Модель Крика і Уотсона - це вторинна структура ДНК, заснована на принципі комплементарності і отримала назву подвійної спіралі. Один виток спіралі складається з 10 нуклеотидів, розмір витка 3,4 нм.
Додаткове структурування в просторі двуспиральной молекули ДНК (у конденсованих хромосомах) з утворенням суперспирали є третинної структурою.
3) .Визначення напрямку передачі інформації - «один ген - один фермент» - Бидл і Татум - 50-і роки. (В даний час - один ген - один поліпептид).
4) .Расшіфровка генетичного коду - Ниренберг, Очоа (До 1964 року розшифровані коди для всіх амінокислот). З'ясування механізму експресії генів у прокаріотів і її регуляції - Франсуа Жакоб і Жан Моно - 50-і роки.
5) .70-ті роки і до теперішнього часу - виявлення особливостей експресії генів у еукаріот. Розвиток генетичної інженерії.
Генетичний код і його властивості
Генетичний код - це система запису інформації про послідовність розташування амінокислот в білках за допомогою послідовності розташування нуклеотидів в молекулі ДНК.
Властивості генетичного коду :
1.Код триплетів - кожна амінокислота зашифрована послідовністю трьох нуклеотидів, званих кодоном або кодоном.
2.Код виродилися - кожна амінокислота кодується більш ніж одним кодоном. (Винятки - амінокислоти метіонін і триптофан. Вони кодуються одним кодоном.)
3.Код універсальний - одні й ті ж триплети кодують одні й ті ж амінокислоти у всіх організмів.
4.Генетіческій код не перекривається - триплети нуклеотидів не перекривають один одного. У деяких вірусів виявлено перекривання, але це виняток.
5.Імеются стартовий і термінальний кодони. Стартовий - єдиний для всіх - АУГ. Термінальний трьох видів - УАГ, УАА, УГА.
Функціонально-генетична класифікація генів
В даний час ген розглядається як одиниця функціонування спадкового матеріалу. Ген - це ділянка молекули ДНК, відповідальний за синтез одного поліпептиду.
Розрізняють три види генів:
структурні;
модулятори;
Регулятори.
Структурні гени несуть інформацію про амінокислотах в білках, ферментах, а також про послідовність нуклеотидів в молекулах р-РНК і т-РНК.
Гени-модулятори впливають на функціонування структурних генів, можуть зміщувати в ту чи іншу сторону процес розвитку ознаки. Вони поділяються на:
1) інгібітори або супресори (епістатічние гени),
2) інтенсифікаторами - можуть підвищувати здатність структурних генів до мутацій,
3) модифікатори - можуть впливати на структурні гени по типу комлементарності.
Гени-регулятори контролюють синтез регуляторних білків, а також час включення різних структурних генів в процесі індивідуального розвитку.
Властивості генів:
1. Дискретність дії - кожен ген діє як самостійна одиниця спадковості.
2. Стабільність - при відсутності мутацій ген передається в ряду поколінь в незміненому вигляді.
3. Специфічність дії - кожен ген впливає на розвиток свого певної ознаки.
4. Плейотропія - здатність одного гена забезпечувати розвиток одночасно декількох ознак (синдром Марфана).
5. Присутність у вигляді двох у диплоїдних і більшого числа алелей у поліплоїдних організмів.
6. Дія гена дозовано, при зміні числа доз гена в організмі змінюється ознака (наприклад, при хворобі Дауна відбувається збільшення до трьох доз генів 21 хромосоми)
Функціональна активність генів або експресія генів
У прокаріотів вона здійснюється в два етапи: транскрипція і трансляція.У еукаріот є ще стадія процесингу.
Експресія генів полягає в синтезі на молекулі ДНК молекули і-РНК, комлементарной їй (або транскрипції - переписування, зчитування біологічної інформації) і подальше її використання для синтезу білка. Одиницею транскрипції в ДНК є транскріптон, що перевищує за розмірами структурні гени. Транскріптон в клітинах еукаріот складається з неінформативної (акцепторной) та інформативною зони. Неінформативна зона починається геном-промотором (ділянка з 80 нуклеотидів), до якого приєднується фермент РНК-полімераза, що каталізує процес зчитування. У прокаріотів один вид РНК-полімерази, у еукаріот три.
За геном промотором знаходяться гени-оператори, які пов'язують регуляторні білки (білки, що включають і припиняють транскрипцію).
Інформативна зона складається з структурних генів, розташованих за генами операторами. Структурні гени еукаріот розділені спейсерами - ділянками ДНК, що не несуть інформації.
Крім того, в складі самих структурних генів є інформаційні ділянки - екзонів і неінформаційних - інтрони. У пркаріот спейсеров, екзонів і інтронів немає.
У кожній фазі життєвого циклу в клітці транскрибується тільки 10% структурних генів, а решта гени не активні, але частина з них може включатися в інших фазах життєвого циклу.
В результаті транскрипції у прокаріот відразу утворюється м-РНК (зріла РНК) і відразу ж починається процес трансляції.
У еукаріот транскрибується велика молекула і-РНК, яка містить всі неінформативні ділянки. Вона називається РНК-попередниця або пре-РНК. Тому за транскрипцією настає процесинг, в результаті, якого руйнуються всі неінформативні ділянки: акцепторная зона, спейсери і інтрони, а що залишилися Екзони зшиваються (сплайсинг).
На етапі процесингу до початкового відрізку утворилася РНК приєднується послідовність нуклеотидів, звана ковпачком, до кінцевого - послідовність залишків аденіну - полі- A.
У процессинге відбувається модифікація нуклеотидів в РНК, наприклад, їх метилювання, гідрування.
І тільки після цих перетворень утворюється зріла м-РНК, яка починається вступної послідовністю, званої лідером, і закінчується кінцевий послідовністю - трейлером. Лідер - вступна послідовність нуклеотидів, комплементарная послідовності в молекулі р-РНК малої субодиниці рибосом, яка забезпечує прикріплення і-РНК до малої субодиниці. Трейлер включає нонсенс-кодон і полі-А послідовність.
Кодовою елементом м-РНК є триплет нуклеотидів, званий кодоном. Кожному кодону відповідає певна амінокислота.
Первинною структурою РНК є, як і в ДНК, лінійна ланцюг полинуклеотидов, в якій мононуклеотиди з'єднані 3`, 5`-фосфоефірную зв'язками.
Вторинна структура РНК - вигнута ланцюг, а третинна являє собою нитку, намотану на котушку, роль котушки грає особливий транспортний білок - інформатор.
Утвориться м-РНК йде в цитоплазму до місця синтезу поліпептиду (білка), тобто до рибосом.
На рибосомах здійснюється процес трансляції. Трансляція - це механізм, за допомогою якого послідовність нуклеотидів в молекулах м-РНК переводиться в специфічну послідовність амінокислот у поліпептидному ланцюзі.
Трансляція складається з трьох стадій:
1. Ініціація - початок синтезу поліпептиду.
2. Елонгація - подовження поліпептиду.
3. Терминация - закінчення синтезу поліпептиду.
На етапі ініціації менша субодиниця рибосоми дізнається стартовий кодон АУГ м-РНК і прикріплюється до неї. АУГ займає першу позицію. Після цього приєднується велика субодиниця рибосоми і в ній починається власне синтез білка. До великої субодиниці підходить т-РНК з амінокислотою.

У молекулі т-РНК одна її частина приєднує амінокислоту, а інша - антикодон, злучається з колоном м-РНК, що визначає цю амінокислоту.

У великій субодиниці є:
1 - аміноацільний ділянку
2 - пептідільний ділянку
У т-РНК, антикодон якої комплементарний кодону АУГ
м-РНК приносить амінокислоту метіанін і зупиняється в аміноацільном ділянці. Стадія ініціації контролюється факторами ініціації. У прокаріотів їх 3, у еукаріот 6-7.
Друга стадія - елонгації починається з переміщення т-РНК з метіанін в пептідільний ділянку, а на її місце приходить інша т-РНК з новою амінокислотою. Між двома амінокислотами утворюється пептидний зв'язок. Тим часом рибосома просувається уздовж м-РНК, на рибосомі виявляється новий кодон, до якого незабаром приєднується своїм антикодоном відповідна т-РНК.
Все це повторюється багато разів, до тих пір, поки рибосома НЕ
дійде до одного з термінальних кодонів (УАА, УАГ, УГА).
Цим кодонам немає антикодону, немає амінокислоти. Настає фаза термінації. До однієї молекулі м-РНК прикріплюється зазвичай багато
рибосом і утворюються полісоми.
У процесі трансляції поряд з факторами ініціації і елонгації важливе значення має фермент, що зв'язує т-РНК з аміноацільним ділянкою - аміноацил т-РНКаза.
Регуляція експресії генів у про- та еукаріот
У прокаріотів вона здійснюється на рівні промотора, оператора
і трансляції.
У еукаріот регуляція відбувається на всіх етапах експресії: транскрипції, процесингу і трансляції.
Детальніше зупинимося на регуляції експресії генів у бактерій.
Регуляція експресії генів у прокаріотів

Схема регуляції транскрипції структурних генів прокаріотів клітини за типом репресії

Схема регуляції транскрипції структурних генів прокаріотів клітини за типом індукції
У 1961 році Жакоб і Моно встановили, що у бактерій під контролем гена-регулятора синтезується білок-репрессор, який регулює активність інших генів. Білок-репрессор зв'язується з геном-оператором і блокує його, внаслідок чого транскрипція (зчитування інформації) стає неможливою і клітина не може синтезувати відповідні білки. Але якщо в клітку проникає будь-якої субстрат, для розщеплення якого необхідно синтезувати ферменти, то відбувається наступне: субстрат приєднується до репрессора і позбавляє його здатності блокувати ген-оператор.
До ДНК приєднуються РНК-синтезують ферменти і починається транскрипція генів. Синтез білка-ферменту припиняється, якщо субстрат повністю розщеплюється, білок-репрессор звільняється і знову блокує гени-оператори. Це приклад регуляції по типу індукції. Також існує регуляція за типом репресії: коли кінцеві продукти біохімічної реакції, з'єднуючись з неактивним білком-репрессором, утворюють комплекс, який блокує роботу гена-оператора.
Антітермінація - полягає в тому, що відбувається ігнорування термінальних кодонів, процес експресії триває.
Регуляція у еукаріот
1) Регуляція генної активності у еукаріот набагато складніше, ніж у бактерій. У еукаріот вона відбувається не тільки на рівні клітини. Існують системи регуляції організму як цілого. Величезну роль в регуляції грають гормони, але регулюють вони процеси синтезу білків лише в клітинах-мішенях. Гормони зв'язуються з білками-рецепторами, розташованими в мембранах таких клітин і включають системи зміни структури клітинних білків. Ті, в свою чергу, можуть впливати як на процеси транскрипції, так і процеси трансляції. Кожен гормон через систему посередників активує свою групу генів. Так адреналін включає синтез ферментів, що розщеплюють глікоген м'язів до глюкози, а інсулін впливає на утворення глікогену з глюкози в печінці.
2) На стадії транскрипції білки-гістони беруть участь в процесах регуляції генної активності у еукаріот. Неодмінна умова - це деконденсація ділянки, де відбувається транскрипція.
3) Регуляція на рівні трансляції спрямована на збереження стабільності м-РНК, а ефективність трансляції здійснюється за допомогою факторів ініціації, елонгації, термінації.
Лекція 12.
мінливість
план
1.Визначення та форми мінливості: модификационная, комбинативная, мутационная.
2.Мутагенние чинники.
3.Классификация мутацій.
4.Устойчівость і способи репарації генетичного матеріалу.
5.Закон гомологічних рядів спадкової мінливості М.І.Вавілова.
Визначення та форми мінливості
Генетика вивчає дві основні властивості живих істот - спадковість і мінливість.
Мінливість - властивість організмів набувати нових ознак і особливості індивідуального розвитку під впливом факторів середовища.
Мінливість - один з найважливіших чинників еволюції, що забезпечують все різноманіття живої природи.
Розрізняють два види мінливості:
1. фенотипическим (ненаследственную, модификационную);
2. генотипічними (спадкову):
а) комбинативную;
б) мутационную.
Модификационная мінливість - форма мінливості, не пов'язана зі зміною генотипу і викликана впливом факторів середовища.
Модификационная мінливість має особливості:
Чи не зачіпає спадкову основу організму і тому модифікації не передаються у спадок, тобто від батьків до потомства,
Зміни спрямовані, відбуваються закономірно, їх можна передбачити,
Мають адаптивне (пристосувальне) значення,
Мають масовий (груповий) і оборотний характер,
Певний фактор зовнішнього середовища викликає подібне зміна у всіх особин даного виду.
Модификационная мінливість має межу. Межі мінливості ознаки, обумовлені генотипом називаються нормою реакції. Вона може бути вузькою, коли ознака змінюється незначно (колір очей), і широкої, коли ознака змінюється в широких межах (зростання, маса тіла).
У медицині часто доводиться встановлювати норму реакції для оцінки max і min кількісних показників (рівень гормонів, ферментів, гемоглобіну та ін.)
Комбинативная мінливість - це спадкова мінливість, обумовлена перекомбинацией наявних генів і хромосом, без зміни структури генів і хромосом (спадкового матеріалу). Цей тип мінливості проявляється вже на стадії утворення статевих клітин.
Джерелами комбинативной мінливості є процеси, що відбуваються в мейозі і в результаті запліднення:
1. Рекомбінація генів при кроссинговере в профазе1 мейозу.
2. Рекомбінація хромосом в ході мейозу (незалежне розбіжність хромосом і хроматид при мейозі)
3. Комбінація хромосом в результаті злиття гамет при заплідненні (хаотичне нагромадження гамет при заплідненні).
Комбинативная мінливість забезпечує генотипическое різноманітність людей, пояснює наявність ознак у дітей і онуків від родичів по батьківській і материнській лінії.
мутационная мінливість - здатність генетичного (спадкового) матеріалу змінюватися і ці зміни успадковуються в потомстві.
В основі мутаційної мінливості лежать мутації.
Мутації - це раптові зміни генетичного матеріалу під впливом середовища і передаються у спадок.
Частота мутацій залежить від виду організму, від віку, від фази онтогенезу, стадії гаметогенезу, може відбуватися в статевих і соматичних клітинах, мати рецесивний і домінантний характер. Наприклад, у людини до 6% гамет несуть мутантні гени.
Процес утворення мутацій називається мутагенезу.
Чинники, що викликають мутації називаються мутагенними.
Мутації спочатку діють на генетичний матеріал особини, а через генотип змінюється і фенотип.
мутагенні фактори
Чинники, що викликають мутації називаються мутагенними факторами (мутагенами) і поділяються на:
1. Фізичні;
2. Хімічні;
3. Біологічні.
До фізичних мутагенних чинників відносяться різні види випромінювань, температура, вологість і ін. Найбільш сильне мутагенну дію надає іонізуюче випромінювання - рентгенівські промені, α-, β-, γ- промені. Вони мають велику проникаючу здатність.
При дії їх на організм вони викликають:
а) іонізацію тканин - утворення вільних радикалів (ОН) або (Н) з води, що знаходиться в тканинах. Ці іони вступають в хімічну взаємодію з ДНК, розщеплюють нуклеїнових кислот і інші органічні речовини;
б) ультрафіолетове випромінювання характеризується меншою енергією, проникає тільки через поверхневі шари шкіри і не викликає іонізацію тканин, але призводить до утворення димерів (хімічні зв'язки між двома піримідинових основ одного ланцюжка, частіше Т-Т). Присутність димарів в ДНК призводить до помилок при її реплікації, порушує зчитування генетичної інформації;
в) розрив ниток веретена поділу;
г) порушення структури генів і хромосом, тобто освіту генних і хромосомних мутацій.
До хімічних мутагенів відносяться:
Природні органічні і неорганічні речовини (нітрити, нітрати, алкалоїди, гормони, ферменти та ін.);
Синтетичні речовини, які раніше не зустрічалися в природі (пестициди, інсектициди, харчові консерванти, лікарські речовини).
Продукти промислової переробки природних сполук - вугілля, нафти.
Механізми їх дії :
а) дезаминирование - відщеплення аміногрупи від молекули амінокислот;
б) пригнічення синтезу нуклеїнових кислот;
в) заміна азотистих основ їх аналогами.
Хімічні мутагени викликають переважно генні мутації і діють в період реплікації ДНК.
До біологічних мутагенів відносяться:
Віруси (грипу, краснухи, кору)
Механізми їх дії:
а) віруси вбудовують свою ДНК в ДНК клітин господаря.
Біологічні мутагени викликають генні і хромосомні мутації.
Класифікація мутацій
Розрізняють такі основні типи мутацій:
1. За способом виникнення їх поділяють на спонтанні та індуковані.
спонтанні- відбуваються під дією природних мутагенних чинників зовнішнього середовища без втручання людини. Вони виникають в умовах природного радіоактивного фону Землі у вигляді космічного випромінювання, радіоактивних елементів на поверхні землі.
індуковані мутації викликаються штучно впливом певних мутагенних чинників.
2. За мутував клітинам мутації підрозділяються на генеративні і соматичні.
генеративні- відбуваються в статевих клітинах, передаються у спадок при статевому розмноженні.
соматичні- відбуваються в соматичних клітинах і передаються тільки тим клітинам, які виникають з цієї соматичної клітини. Вони не передаються у спадок.
3. За впливом на організм:
Негативні мутації - летальні (несумісні з життям); напівлетальні (знижують життєздатність організму); нейтральні (які не впливають на процеси життєдіяльності); позитивні (підвищують життєздатність). Позитивні мутації виникають рідко, але мають велике значення для прогресивної еволюції.
4. По змін генетичного матеріалу мутації підрозділяються на геномні, хромосомні і генні.
геномні мутації - це мутації, викликані зміною числа хромосом. Можуть з'являтися зайві гомологічні хромосоми. У хромосомному наборі на місці двох гомологічних хромосом виявляються три - це трисомія. У разі моносомии спостерігається втрата однієї хромосоми з пари. При полиплоидии відбувається кратне гаплоидному збільшення числа хромосом. Ще один варіант геномної мутації - Гаплоїдія, при якій залишається тільки одна хромосома з кожної пари.
хромосомнімутації пов'язані з порушенням структури хромосом. До таких мутацій відносяться втрати ділянок хромосом (делеції), додавання ділянок (Дуплікація) і поворот ділянки хромосом на 180 ° (інверсія).
геннімутації, при яких зміни відбуваються на рівні окремих генів, тобто ділянок молекули ДНК. Це може бути втрата нуклеотидів, заміна одного підстави на інше, перестановка нуклеотидів або додавання нових.
Стійкість і репарація генетичного матеріалу
Стійкість до змін генетичного матеріалу забезпечується:
1. диплоїдний набір хромосом.
2. Подвійний спіраллю ДНК.
3. виродження (надмірністю) генетичного коду
4. Повтором деяких генів.
5. репарації порушень структури ДНК
Наявність механізмів репарації - обов'язкова умова існування біологічних істот.
Репарація генетичного матеріалу – це процес, що забезпечує відновлення пошкодженої структури молекули ДНК.
У ДНК клітини щодня відбувається безліч випадкових змін.
Більшість ефективно виправляються (репаруючу) за допомогою спеціальних ферментних систем.
Вперше репарація молекули ДНК була встановлена в 1948 році. А в 1962 році був описаний один із способів репарації - світлова репарація або фотореактивації.
Було встановлено, що при ультрафіолетовому опроміненні вірусів-фагів, бактерій і найпростіших спостерігається різке зниження їх життєдіяльності, навіть загибель.
Якщо впливати на них видимим світлом, то виживаність їх значно збільшується.
Виявилося, що під дією ультрафіолету в молекулі ДНК утворюються димери (хімічні зв'язки між двома підставами одного ланцюжка, частіше Т-Т), освіту димарів перешкоджає зчитуванню інформації.
Видиме світло активує ферменти, що руйнують димери.
Другий спосіб репарації - темновая репарація, була вивчена в 50-і роки ХХ століття.
Темнова репараціяпротікає в чотири стадії за участю чотирьох груп ферментів. Ферменти утворилися в ході еволюції і спрямовані на підтримку стабільності генетичної інформації клітини.
1. Фермент ендонуклеаза знаходить пошкоджену ділянку і поруч з ним розриває нитку ДНК.
2. Фермент ектонуклеаза «вирізає» (видаляє) пошкоджену ділянку.
3. ДНК-полімераза за принципом комплементарності синтезує фрагмент ДНК на місці зруйнованого.
4. лігаза «зшиває» синтезований фрагмент з основною ниткою ДНК.
Доведено можливість репарації ДНК при пошкодженні обох її ниток. При цьому інформація може бути отримана з і-РНК (фермент ревертаза).
Закон гомологічних рядів спадкової мінливості М.І.Вавілова
Відомо, що мутирование відбувається в різних напрямках. Однак, це різноманіття підпорядковується певної закономірності, виявленої в 1920 році М. І. Вавілов. Він сформулював закон гомологічних рядів спадкової мінливості: «Види і пологи, генетично близькі, характеризуються подібними рядами спадкової мінливості з такою правильністю, що знаючи ряд форм у межах одного виду, можна передбачати існування паралельних форм у інших видів і родів».
Можна сказати, що у споріднених видів, що мають спільне походження виникають подібні мутації.
Одні і ті ж хвороби зустрічаються у тварин і у людини. Так, наприклад, альбінізм спостерігається у всіх класах хребетних тварин і людини. Брахидактилия (короткопалость) відзначена у щурів, овець, собак, людини. М'язова дистрофія - у мишей, щурів, коней, людини.
Закон гомологічних рядів дозволяє передбачити можливість появи мутацій, які можуть бути використані в селекції для створення нових цінних для господарства форм.
Методи вивчення генетики людини
план
1.Генеалогіческій метод.
2.Блізнецовий метод.
3.Метод дерматоглифики.
4.Цітогенетіческій метод.
5.Методи гібридизації соматичних клітин.
6.Онтогенетіческій метод.
7.Популяціонно-статистичний метод.
8.Метод моделювання.
9.Іммунологіческій метод.
10.Біохіміческій метод.
генеалогічний метод
Типи успадкування і форми прояву генетичних задатків у людини дуже різноманітні і для диференціації між ними потрібні спеціальні методи аналізу, в першу чергу - генеалогічний, запропонований Ф.Гальтона.
Генеалогічний метод або вивчення родоводів передбачає простеження ознаки в сім'ї або роду із зазначенням типу родинних зв'язків між членами родоводу. У медичній генетиці цей метод зазвичай називають клініко-генеалогічним, оскільки мова йде про спостереження патологічних ознак з допомогою прийомів клінічного обстеження. Генеалогічний метод відноситься до найбільш універсальним методам в генетиці людини. Він широко застосовується при вирішенні теоретичних і практичних проблем:
1) для встановлення спадкового характеру ознаки,
2) при визначенні типу успадкування та пенетрантности генотипу,
3) виявлення зчеплення генів і картування хромосом,
4) при вивченні інтенсивності мутаційного процесу,
5) при розшифровці механізмів взаємодії генів,
6) при медико-генетичному консультуванні.
Суть генеалогічного методу зводиться до з'ясування родинних зв'язків і простежуванню ознаки серед близьких і далеких прямих і непрямих родичів. Технічно він складається з двох етапів: складання родоводів і генеалогічного аналізу.
складання родоводу
Збір відомостей про сім'ю починається з пробанда, яким називається особа, першим потрапило в поле зору дослідника.
Діти однієї батьківської пари (рідні брати і сестри) називаються сибсами. Сім'єю у вузькому сенсі, або ядерної сім'єю, називають батьківську пару і їх дітей. Більш широке коло кровних родичів краще позначати терміном «рід». Чим більше поколінь втягується в родовід, тим вона ширше. Це тягне за собою неточність отриманих відомостей і, отже, неточність родоводу в цілому. Часто люди не знають навіть числа своїх двоюрідних братів і сестер, не кажучи вже про якісь ознаки у них і їх дітей.
Для наочності готують графічне зображення родоводу. Для цього зазвичай користуються стандартними символами. Якщо розглянутих ознак в родоводі багато, то можна вдаватися до літерним або штриховим відмінностей всередині символів. Схема родоводу обов'язково супроводжується описом позначень під малюнком - легендою, що виключає можливість неправильних тлумачень.
генеалогічний аналіз
Метою генеалогічного аналізу є встановлення генетичних закономірностей.
1 етап – встановлення спадкового характеру ознаки. Якщо в родоводі зустрічається один і той самий ознака кілька разів, то можна думати про спадкової його природі. Однак треба перш за все виключити можливість екзогенного накопичення випадків в родині або роду. Наприклад, якщо один і той же патогенний фактор діяв на жінку під час всіх вагітностей, то у неї можуть народитися декілька дітей з однаковими аномаліями. Або ж якийсь фактор діяв на багатьох членів сім'ї, необхідно звірити дію подібних зовнішніх чинників. За допомогою генеалогічного методу були описані всі спадкові хвороби.
2 етап – встановлення типу успадкування та пенетрантности гена. Для цього використовують принципи як генетичного аналізу, так і статистичні методи обробки даних з родоводом.
3 етап – визначення груп зчеплення і картування хромосом, до недавнього часу грунтується тільки на генеалогічному методі. З'ясовують зчеплені ознаки і процес кросинговеру. Цьому сприяють розроблені математичні методи.
4 етап – вивчення мутаційного процесу. Він застосовується в трьох напрямках: при вивченні механізмів виникнення мутацій, інтенсивності мутаційного процесу і факторів, що викликають мутації. Особливо широко генеалогічний метод застосовується при вивченні спонтанних мутацій, коли треба розрізняти «спорадично» виникли випадки від «сімейних».
5 етап – аналіз взаємодії генів в клінічній генетиці був зроблений С.Н.Давіденковим (1934, 1947) з аналізу поліморфізму захворювань нервової системи.
6 етап – в медико-генетичному консультуванні для складання прогнозу без генеалогічного методу обійтися не можна. З'ясовують гомо- чи гетерозиготность батьків і розглядають ймовірність народження дітей з тими чи іншими ознаками.
Близнюковий метод дослідження
Дослідження близнюків - один з основних методів генетики людини. Існують однояйцеві близнюки, що виникають з однієї яйцеклітини, заплідненої одним сперматозоїдом. Виникають вони через поділ зиготи на два генетично ідентичних один одному і завжди одностатевих зародка.
Разнояйцовие близнюки розвиваються з різних яйцеклітин, запліднених різними сперматозоїдами. Генетично вони розрізняються як брати і сестри одних батьків.
За допомогою близнецового методу можна вивчити:
1) Роль спадковості і середовища у формуванні фізіологічних і патологічних особливостей організму. Зокрема, вивчення спадкової передачі людьми деяких хвороб. Вивчення експресивності і пенетрантности генів, що викликають спадкові захворювання.
2) Конкретні чинники, які посилюють або послаблюють вплив зовнішнього середовища.
3) Кореляцію ознак і функцій.
Особливо важлива роль близнецового методу у вивченні проблеми «генотип і середовище».
Порівнюють зазвичай три групи близнюків: ДБ в однакових умовах, ПРО в однакових умовах, ПРО в різних умовах.
При вивченні близнюків визначають частоту, ступінь збігу (конкордантности) тих чи інших ознак.
При вивченні ролі спадковості в походженні тієї чи іншої ознаки роблять розрахунок за формулою К.Хольцінгера.
Коефіцієнт успадкованого - Н
Н =% подібності ОБ -% подібності РБ
100 -% подібності РБ
При Н = 1 вся мінливість в популяції обумовлена спадковістю.
При Н = 0 вся мінливість викликана середовищні фактори. Вплив середовища З виражається формулою: С = 100% - Н, де Н - коефіцієнт успадкованого. Наприклад, конкордантность монозиготних (однояйцевих) близнюків 3%. Тоді Н =
67 – 3 = 64 = 0,7 або 70%. С = 100 - 70 = 30%
Отже, дана ознака на 70% обумовлений спадковістю, а на 30% - впливом факторів зовнішнього середовища.
Інший приклад. Групи крові за системою АВО у ПРО = 100%, тобто повністю залежить від спадковості.
Частота збігу груп крові і деяких захворювань у близнюків (в%)
|
Ознаки або хвороби | |||
|
групи крові АВО | |||
|
шизофренія | |||
|
епілепсія | |||
|
Природжений стеноз воротаря |
метод дерматогліфіки
Ето наука, що вивчає спадкову обумовленість малюнків, які утворюють лінії шкіри на кінчиках пальців, долонях і підошвах людини.
Виявилося, що у кожного народу, у кожної раси, у кожної людини малюнки мають свої особливості, і на долонях вони строго індивідуальні. На це вперше звернув увагу Ф. Гальтон, який запропонував англійської кримінальної поліції за відбитками пальців ідентифікувати злочинців.
Дерматогліфічні дослідження мають важливе значення в криміналістиці, в визначенні зиготности близнюків, в діагностиці ряду спадкових захворювань, а так само в окремих випадках спірного батьківства.
Долонний рельєф дуже складний. У ньому виділяють ряд полів, подушечок і долонних ліній. Подушечок на долоні 11, їх ділять на 3 групи:
1) п'ять кінцевих (еплікальних) подушечок на кінцевих фалангах пальців.
2) чотири міжпальцевих подушечки, розташовуються проти міжпальцевих проміжків.
3) дві долоні проксимальні подушечки тенар і гіпотенара. У підстави великого пальця - тенар, у протилежного краю долоні-гіпотенара.
На найбільш піднесених частинах подушечок помітні шкірні гребінці. Це лінійні потовщення епідермісу, які представляють собою модифіковані лусочки шкіри. Шкірні гребінці йдуть потоками, як на долонях, так і на пальцевих подушечках. Точки зустрічі цих потоків утворюють трирадіус або дельти.
Гребешкова візерунки зазвичай вивчають під лупою. Відбитки візерунків, за допомогою друкарської фарби, роблять на чистій білій, краще крейдованому, папері або целофані. Як на кінчиках пальців, так і на долонних підвищеннях можуть спостерігатися різні папілярні візерунки у вигляді завитків, петель і дуг, відкритих в ульпарную або радіальну боку. На Тенар і гіпотенара частіше бувають дуги. На середньої і основний фалангах пальців Гребешкова лінії йдуть поперек пальців, утворюючи різні візерунки - прямі, серповидні, хвилеподібні, дугоподібні і їх поєднання. В середньому на одному пальці бувають 15-20 гребінців.
Малюнок долоні.

1 - поперечна проксимальная борозна, лінія притиснення 4 пальців
2 - поперечна середня борозна, лінія притиснення 3 пальців
3 - поперечна дистальная борозна, лінія притиснення 2 пальців
4 - борозна великого пальця
5 - поздовжня серединна борозна від зап'ястя до основи 3го пальця
6 - поздовжня проміжна борозна від зап'ястя до основи 4го пальця
7 - поздовжня ульнарная борозна, від зап'ястя до основи 5го пальця

1 - синдром Патау
2 - синдром Дауна
3 - синдром Шерешевського-Тернера
4 - норма
5 - синдром Клайнфельтера
При вивченні кожного рельєфу долоні досліджують:
1) Хід головних долонних ліній А, В, С, Д 1,2,3,4,5,6,7.
2) Долонні візерунки на Тенар і гіпотенара.
3) Пальцеві візерунки (форму візерунків, гребневий рахунок)
4) Осьові трирадіус.
Аналогічні дослідження проводять і на підошвах ніг. Напрямок головної долонної лінії Д у батьків і їх дітей однакове.
Вивчення хворих з хромосомними хворобами (хвороба Дауна, синдром Клайнфельтера) показало, що у них змінюється не тільки малюнок пальцевих і долонних візерунків, а й характер основних згинальних борозен на шкірі долонь.
Дещо меншу вираженість мають дерматогліфічні відхилення у хворих з такими дефектами розвитку, як вроджені пороки серця і магістральних судин, незарощення м'якого і твердого неба, верхньої губи і т.д.
Встановлено зміни в характері пальцевих і долонних візерунків при витівці, шизофренії, цукровому діабеті, раку, ревматизмі, поліомієліті та інших захворюваннях.
цитогенетичний метод
Цей метод дозволяє за допомогою мікроскопа досліджувати структури клітини - хромосоми. За допомогою методу мікроскопії вивчений каріотип організму людини (хромосомний набір клітин організму). Встановлено, що багато захворювань і дефекти розвитку пов'язані з порушенням числа хромосом і їх будови. Цей метод дозволяє вивчити також дію мутагенів на склад і будова хромосом. Цитогенетичний метод пов'язаний з тимчасовими культурами тканин (зазвичай лейкоцитів) і отриманням метафазних ядер з укороченими, потовщеними хромосомами, розподіл яких зупиняють на стадії метафазної пластинки колхицином. Якщо в каріотипі вивчаються статеві хромосоми, то цей метод дозволяє досліджувати статевої хроматин в соматичних клітинах.
Гібридизація соматичних клітин
Гібридні клітини мають певні властивості, що дозволяють визначати локалізацію гена або зчеплення гена. Втрата хромосом людини з деяких типів гібридних клітин дозволяє отримувати клони з відсутністю певної хромосоми. Найбільш споживані гібриди соматичних клітин людина - миша.
Відстеження за наявністю біохімічного генетичного маркера в гібридних клонах в міру елімінації хромосом людини може привести до виявлення локалізації гена, якщо ознака зникає з клітин, як тільки вони міняються певними хромосомами. Цитогенетичний аналіз великого числа клонів і зіставлення результатів з присутністю великої кількості генетичних маркерів дозволяє помітити зчеплені гени і їх локалізацію. Додатково використовують інформацію, при використанні клонів від інвалідів з транслокаціями і іншими хромосомними аномаліями.
Цим методом було встановлено локалізація гена фосфогліцераткинази в довгому плечі Х-хромосоми, тобто місце гібридних клітин дозволяє встановити:
1) локалізацію гена
2) зчеплення генів
3) картування хромосом
Понад 160 локусів визначені за допомогою методу гібридних соматичних клітин.
онтогенетический метод
Дозволяє вивчити закономірності прояву якої-небудь ознаки або захворювання в процесі індивідуального розвитку. Виділяють кілька періодів розвитку людини. Антенатальний (розвиток до народження) і постнатальний. Більшість ознак людини формуються в фазу морфогенезу антенатального періоду. У фазу морфогенезу постнатального періоду закінчується формування кори головного мозку і деяких інших тканин і органів, формується імунологічна система організму, яка досягає найвищого розвитку через 5-7 років після народження дитини. У постморфогенетіческій період розвиваються вторинні статеві ознаки.
У морфогенетичний період зміна активності генів відбувається за двома типами:
1) включення і виключення генів
2) посилення та ослаблення дії генів
У постморфогенетіческій період розвитку перший тип зміни активності генів майже відсутня, відбувається лише невелике включення окремих генів - наприклад, генів, що визначають вторинні статеві ознаки, розвиток деяких спадкових захворювань. Вимкнення же генів в цьому періоді більш значне. Репресує активність багатьох генів, пов'язаних з виробленням меланіну (в результаті відбувається посивіння), а також генів, пов'язаних з виробленням γ-глобулінів (підвищується сприйнятливість до захворювань). Придушуються багато генів в клітинах нервової системи, м'язових клітинах і т.д.
Репресія генів здійснюється на рівні транскрипції, трансляції, посттрансляціі. Однак основний тип зміни активності генів на цьому етапі - посилення і ослаблення дії генів. Може змінюватися домінування генів, що викликає зміна зовнішніх ознак, особливо в період статевого дозрівання. Змінюється співвідношення статевих гормонів і відповідно ознаки статі. Репресивні гени з віком можуть надавати великий вплив на розвиток тієї чи іншої ознаки. Наприклад, ген фенілкетонурії в гетерозиготному стані змінює психіку людини.
Популяційно-статистичний метод дослідження
Являє собою метод математичного підрахунку тих чи інших генів і відповідних ознак в певних популяціях. Теоретичною основою даного методу є закон Харді-Вайнберга.
Цим методом встановлено, що всі гени людської популяції по частоті можна розділити на 2 категорії:
1) мають універсальне поширення, до складу яких входить більшість генів. Наприклад, ген дальтонізму, наявний у 7% чоловіків і більш ніж у 13% жінок. Ген ідіотії, зустрічається у населення Європи з частотою 4 на 10 000 населення.
2) гени, що зустрічаються переважно в певних районах. Наприклад, ген серповидно-клітинної анемії поширений в країнах, де лютує малярія. Ген вродженого вивиху стегна, що має високу концентрацію у аборигенів північного сходу нашої країни.
метод моделювання
Закон гомологічних рядів М. І. Вавилова (види і роди генетично близькі володіють схожими рядами спадкової мінливості) дозволяє з певними обмеженнями екстраполювати експериментальні дані на людину.
Біологічна модель спадкового захворювання на тваринному часто є більш зручною для дослідження, ніж хвора людина. Виявилося, що у тварин є близько 1300 спадкових хвороб, так само, як у людини. Наприклад, у мишей - 100, у крокодилів - 50, у щурів - 30. на моделі гемофілії А і В у собак показано, що вона обумовлена рецесивним геном, розташованим в Х-хромосомі.
Моделювання м'язової дистрофії у мишей, хом'яків і курей дало можливість зрозуміти патогенетичну сутність цього захворювання. Було встановлено, що при цьому захворюванні уражається нервова система, а безпосередньо м'язові волокна.
Початкові механізми галактоземии були з'ясовані на моделі кишкової палички. І у людини, і у бактерій нездатність засвоювати галактозу викликана однаковим спадковим дефектом - відсутністю активного ферменту - галактоза-1-фосфатілуріділтрансферази.
Імунологічний метод дослідження
Цей метод заснований на вивченні антигенного складу клітин і рідин людського організму - крові, слини, шлункового соку і т.п. Найчастіше досліджують антигени формених елементів крові: еритроцитів, лейкоцитів, тромбоцитів, а також білків крові. Різні види антигенів еритроцитів утворюють системи груп крові.
На початку ХХ століття К.Ландштейнером і Я.Янскій показали, що в залежності від характеру реакцій між еритроцитами і плазмою крові, всіх людей можна розділити на 4 групи. Надалі було доведено, що реакції ці відбуваються між білковими речовинами еритроцитів, які були названі агглютіно-генами, і білками сироватки крові, які були названі агглютининами.
Групи крові визначаються антигенами, що містять ліпідну і білкові фракції, і які знаходяться на поверхні еритроцитів. Білкова частина антигену контролюється геном, який працює на ранніх етапах розвитку еритроцита. Антигени специфічні для кожної групи крові.
Всього зараз відомо 14 систем еритроцитарних груп крові, в які входять більше 100 різних антигенів. В системі груп крові АВО на поверхні еритроцитів формується два антигену під контролем генних алелей I а , I в .
Бернштейн в 1925 році показав, що є третя аллель I про , Яка не контролює синтез антигену. Таким чином, в системі АВО груп крові існує три алелі, але у кожної людини є тільки два з них. Якщо розписати можливі чоловічі і жіночі гамети в решітці Пеннета, то можна простежити, які можливі комбінації груп крові будуть у нащадків.
Групи крові АВО у нащадків залежно від груп крові у батьків
Імунологічні методи застосовують для обстеження пацієнтів та їхніх родичів при підозрі на іммунодефецітние стану (агаммаглобулинемия, дісгаммаглобулінемія, атаксія-телеангіектазії та інші), при підозрі на антигенну несумісність матері і плоду, при пересадці органів і тканин, при встановленні істинного споріднення, у випадках медико- генетичного консультування, при необхідності вивчення генетичних маркерів при діагностиці зчеплення генів або при визначенні спадкової схильності до захворювань, п ри встановленні зиготности близнюків.
Визначення групової приналежності крові має практичне значення в різноманітних генетичних дослідженнях:
1) при встановленні зиготних близнюків
2) при встановленні зчеплення генів.
3) в судово-медичній експертизі у випадках спірного батьківства чи материнства. Відомо, що у дитини не могли з'явиться антигени, яких немає у батьків.
Система груп крові М була відкрита К.Ландштейнером і І.Левіним в 1927 році (в цій групі антитіла до відповідних антигенів не продукує). У системі є два алелі M, N.
Гени, що визначають фактор М і N, є Кодомінантність, тобто якщо зустрічаються разом, то обидва і проявляються. Таким чином, існують гомозиготні генотипи MM і NN, і гетерозиготні MN. У популяціях європейців генотипи ММ зустрічаються приблизно в 36%, NN - в 16%, MN - в 48%.
А гени відповідно:
М = 36 + 48 = 60%
Резус фактор
Як показали дослідження вчених, 85% європейців мають еритроцитарний антиген, загальний з антигеном мавп виду макака-резус. У 15% людей резус-антигенів на поверхні еритроцитів немає.
Система групових резус-антигенів дуже складна. Передбачається, що резус-антигени контролюються трьома тісно зчепленими локусами С, Д і Е в двох хромосомах і успадковуються домінантно. Тому можливі по кожному локусу три генотипу: гомозиготні резус-позитивні, гетерозиготні резус-позитивні і гомозиготні резус-негативні.
Найбільш імуногенним є антиген Д . Антигени С і Е є менш активні.
У 1962 році було встановлено наявність еритроцитарного ізоантигени Х д, що передається через статеву Х-хромосому. З цього антигену всіх людей можна розділити на Х д-позитивних і Х д-негативних. Серед Х д-позитивних жінок зустрічається 88%, а серед чоловіків - 66%. Якщо обоє батьків Х д-негативних, то все їх діти (як дівчатка, так і хлопчики) будуть Х д-негативних. Якщо батько ХД-позитивний, а мати Х д-негативних, то їх дочки будуть Х д позитивними, а сини Х д-негативних. Якщо мати Х д-позитивний, а батько Х д -отріцателен, то їхні сини будуть Х д позитивними тобто тип спадкування «хрест-навхрест». Дочки ж можуть бути як ХД-позитивними, так і Х д-негативних, в залежності від гомозиготности матері. Ген Х д - групи локалізована в короткому плечі Х-хромосоми. Система Х д використовується для вивчення анеуплоїдій (аномального числа Х-хромосом у дитини з трисомія Х, синдрому Клайнфельтера, синдрому Шерешевського-Тернера та ін.). Передбачається, що Х д -несовместімость матері і плоду (мати Х д-негативних, а плід Х д-позитивний) призводить до зменшення частоти народження дівчаток.
біохімічний метод
Дозволяє, з одного боку, вивчити кількість ДНК в клітинах людини в нормі та патології, з іншого - визначати спадкові дефекти обміну речовин за допомогою:
1) визначення аномального білка (структурних білків або ферментів), які утворюються в результаті біохімічних реакцій;
2) визначення проміжних продуктів обміну, які з'являються внаслідок генетичного блоку прямої реакції обміну.
Наприклад, при фенілкетонурії амінокислота фенілаланін чи не перетворюється в тирозин. Відбувається збільшення її концентрації в крові і зменшення концентрації тирозину. Фенілаланін при цьому перетворюється в фенілпіровиноградну кислоту і її похідні - фенілмолочная, фенілоцтової і фенілацетілглутаміновую.
Ці сполуки виявляють в сечі хворого за допомогою хлорного заліза FeCl 3 або 2,4 - дінітрофенілгідразіна.
Одним з явищ, видоизменяющих класичне розщеплення за фенотипом 9: 3: 3: 1 при дигибридном схрещуванні, є неповне домінування.
На прикладі моногибридного схрещування у нічної красуні (Mirabilis jalapa) та інших об'єктів було показано, що при неповному домінуванні однієї з алелей гена гібрид першого покоління виявляється проміжним по прояву альтернативних ознак. У потомстві такого гібрида в F 2 відбувається розщеплення за фенотипом у співвідношенні 1: 2: 1, що відповідає розщеплення по генотипу. Той факт, що в даному випадку гетерозиготное потомство відрізняється по зовнішнім виглядом від гомозиготного, дає можливість зробити висновок, що домінування є результат взаємодії пари алелей, що визначають розвиток даної ознаки. Інакше кажучи, домінування відображає переважання однієї алелі гена над іншою (наприклад, А над а, В над b і т. Д.). Очевидно, що така взаємодія членів алельних пари не завжди має своїм результатом повне переважання, т. Е. Повне домінування.
Строго кажучи, ймовірно, генів з повним домінуванням взагалі не існує, так як ген, -як ми переконаємося в подальшому, визначає не один який-небудь ознака або властивість, а впливає на цілий ряд ознак. Тому, кажучи про домінування або рецесивним якогось гена, ми умовно простежуємо лише одну зі сторін його дії - розвиток однієї ознаки.
У дигибридном схрещуванні при неповному домінуванні двох пар алелей розщеплення за фенотипом у F 2 буде відповідати формулі розщеплення (1: 2: 1) 2, або 1: 2: 2: 4: 1: 2: 1: 2: 1. Таким чином, ми будемо мати в F 2. 9 фенотипических класів, що відповідає числу генотипических класів - 3 2. Наведемо два приклади дигибридного схрещування при неповному домінуванні по двох парах алелей у тварини (курка) і рослини (суниця).
У курей відомий ген, що викликає сильну курчавость оперення. Аллель Ã цього гена неповно домінує над своїм нормальним аллелью а, визначальною нормальну будову пера. Є й інша алельних пара, яка перебуває в іншій парі гомологічних хромосом, яка викликає так звану розбризканої забарвлення оперення - чорні цятки на білому тлі. Це фарбування контролюється в повному обсязі домінантною аллель Ḃ. Рецессивная аллель b - визначає чорне забарвлення. При схрещуванні курки з сильною курчавістю і чорним оперенням ÃÃbb з півнем, що має розбризканої забарвлення і нормальне оперення ааḂḂ, гібриди першого покоління ÃаḂb виявляються проміжними по вираженості цих ознак: у них проявляється слабка курчавость і блакитне забарвлення. У другому поколінні такого дигибридного схрещування розщеплення за фенотипом спостерігається не в відношенні 9: 3: 3: 1, а в відношенні 1: 2: 2: 4: 1: 2: 1: 2: 1.
Інший приклад можна привести з дослідів Т. С. Фадєєвої, проведених з аналізу успадкування ознак забарвлення ягоди і форми чашечки у суниці (Fragaria vesca). Червоне забарвлення ягоди визначається гомозиготних станом аллели Ã, біла - аллели а. Інша алельних пара в гомозиготному стані визначає нормальну будову чашечки ḂḂ, а рецессивное гомозиготний стан гена bb призводить до утворення листоподібною чашечки.
Гібридні рослини першого покоління ÃаḂb мають рожеві ягоди і проміжне будова чашечки. Від самозапилення таких форм у другому поколінні розвиваються рослини, які становлять 9 різних фенотипічних класів, що відповідають 9 класів генотипів. Статистично підтверджується збіг теоретично очікуваного розщеплення за фенотипом зі спостережуваним в досвіді: χ 2 = 11,37, 0,20\u003e Р\u003e 0,10.
Найчастіше зустрічаються такі випадки, коли в дигибридном схрещуванні має місце неповне домінування лише в одній з двох пар алелей. Класичним прикладом в цьому відношенні є розщеплення в F 2 за ознаками забарвлення і форми квітки у лев'ячого зіва (Antirrhinum majus). Червоне забарвлення квітки у лев'ячого зіва визначається домінантною аллель з неповним домінуванням її над рецесивною аллель, що обумовлює біле забарвлення. Інша пара ознак стосується форми квітки: нормальна - домінантна і пілорична - рецесивна.

В даному дигибридном схрещуванні розщеплення в F 2 дає 6 фенотипических класів: 3: 6: 1: 2: 3: 1, причому червоні квітки будуть тільки в разі гомозиготного стану домінантною аллели гена забарвлення, рожеві - гетерозиготного її стану, а білі - гомозиготного стану рецессивной аллели цього гена. За формою квітки проміжних форм не буде, так як гетерозиготні і гомозиготні домінантні форми тут не відрізняються один від одного.
Отже, неповне домінування по одній або обом парам алелів генів в дигибридном схрещуванні видозмінює розщеплення в по фенотипу. При повному домінуванні в F 2 буде 4 фенотипических класу, при неповному домінуванні в одній з алельних пар генів - 6 і при неповному домінуванні в обох алельних парах - 9. В останньому випадку розщеплення за фенотипом повністю збігається з розщепленням за генотипом.
Слід ще раз повторити положення, висловлене раніше, що неповне домінування хоча і ускладнює картину менделевского розщеплення в дигибридном схрещуванні, але ні в якій мірі не суперечить законам Менделя, а підтверджує їх. Явище неповного домінування розкриває новий зміст законів успадкування. Якщо погодитися з тим, що ознака обумовлена геном і характер розщеплення в F 2 визначається поведінкою хромосом в мейозі, то особливості розщеплення при неповному домінуванні по-новому розкривають дію гена і характер його прояву. Характер прояву гена також може видозмінювати розщеплення за фенотипом.
Отже, явище незалежного розщеплення при може здійснюватися за таких умов:
- знаходження враховуються генів в негомологічних хромосомах; число їх при цьому не може перевищувати гаплоидное число хромосом у даного виду;
- равновероятное освіту гамет всіх сортів на основі випадкового розбіжності хромосом в мейозі;
- равновероятное дозрівання гамет всіх типів;
- равновероятности зустріч гамет при заплідненні;
- равновероятности виживаність зигот і дорослих організмів;
- відносна стабільність розвитку досліджуваних ознак. Але всі ці умови складаються під впливом різних, постійно і тимчасово діючих факторів. В силу цього вони можуть змінюватися, змінюючи прояв закономірностей успадкування.
Якщо ви знайшли помилку, будь ласка, виділіть фрагмент тексту і натисніть Ctrl + Enter.