Čo je recesívny gén? Recesívne gény
Dedičnosť -schopnosť organizácií opakovať podobné znaky medzi generáciami. Vzhľadom na dedičnosť majú rodičia a potomkovia podobný vzhľad, telesnú štruktúru, metabolizmus. V dôsledku toho sa každý druh reprodukuje z generácie na generáciu.
Dedičnosť je výsledkom kombinácie génov.
gény - tieto sú biochemické funkčné prvky chromozómov, ktoré určujú potenciálny pohlavie a iné príznaky embrya.
chromozóm (Starogréčtina -. Farba a telo) - nukleoproteinové štruktúry v jadre eukaryotických buniek (buniek, ktoré obsahujú jadro), ktoré sa stanú ľahko viditeľné v určitých fázach bunkového cyklu (počas mitózy alebo meiózy). Chromozómy sú vysoký stupeň kondenzácie chromatínu, ktorý je neustále prítomný v bunkovom jadre. Termín bol pôvodne navrhnutý tak, aby odkazoval na štruktúry detegované v eukaryotických bunkách, ale v posledných desaťročiach sa čoraz viac hovorí o bakteriálnych chromozómoch. Tieto chromozómy sú sústredené na väčšinu dedičných informácií.
Chromozómy sú reprezentované dvojicami v jadre každého spermií a každého vajíčka. Osoba má 23 párov alebo 46 chromozómov, s jedným párom nazývaným pohlavné chromozómy, pretože určujú pohlavie budúceho organizmu. Ženské bunky nesú dva X chromozómykdežto samčekové bunky obsahujú jeden X a jeden Y chromozóm, Y chromozóm je menší a obsahuje menej génov na povrchu ako X chromozóm. V dôsledku rozdelenia buniek vo vaječníkoch všetky vajcia obsahujú jeden X chromozóm, zatiaľ čo bunkové delenie v semenníkoch vedie k skutočnosti, že polovica spermií obsahuje X chromozóm a druhá polovica obsahuje chromozóm Y. Napríklad v polovici prípadov sa ľudské vajíčko oplodňuje spermie nesúcou chromozóm X a v druhej polovici spermií nesúci chromozóm Y, takže polovica detí narodených v populácii by mala byť mužská a polovica by mala byť samica.
Dominantný gén(dominantný gén) - gén exprimovaný vo fenotype bez ohľadu na prítomnosť ďalšej alely tohto génu v genóme. Prítomnosť, ktorá zabezpečuje prejav znaku, ktorý určuje, bez ohľadu na to, či je iný gén toho istého páru dominantný alebo recesívny.
odstúpenieexplicitný gén(ustupujúce) - genetické informácie, ktoré môžu byť potlačené expozíciou dominantný gén, Recesívny gén je schopný zabezpečiť prejav vlastnosti, ktorú určí iba vtedy, ak je spárovaný so zodpovedajúcim recesívnym génom. Ak je spárovaný s dominantným génom, potom sa nezjavuje, pretože dominantný gén to potláča. Prezentované vlastnosti recesívne gény, sa objavujú v fenotype potomstva len vtedy, ak majú obaja rodičia recesívny gén.
Ak sú obidve alely v páre úplne totožné (napríklad OO, AA), potom sa takýto genotyp a jeho majiteľ nazývajú homozygotná, a ak sú tieto alely iné (povedzme, AO) - heterozygotná, Je známe, že ak homozygotné genotypy OO a AA predurčujú prvú a druhú krvnú skupinu, potom vlastníci heterozygotného genotypu AO budú tiež mať druhú krvnú skupinu. To znamená, že v takejto kombinácii sa prejavuje účinok génu A a účinok génu O, teda génu A dominantou, a gén O vo vzťahu k nemu ustupujúce (slovo "recesívne" znamená miznutie). Preto dominantné gény vyvíjajú svoje pôsobenie v homozygotnom aj heterozygotnom stave a recesívne gény sa môžu prejaviť iba v homozygotnom stave a nedávajú vonkajšie prejavy u heterozygotných ľudí.
Súbor dedičných faktorov tela (génov) genotypu, Kombinácia všetkých vlastností a vlastností organizmu, ktoré sú výsledkom interakcie genotypu a vonkajšieho prostredia, fenotypom, To znamená, že fenotyp je odrazom genotypu v kombinácii s prostredím. Hranice, v ktorých prostredie ovplyvňuje prejav genotypu, sa nazýva reakčná norma.
Zdroje informácií:
Základy genetiky a chovu psovGenetika je extrémne ťažká veda. Existuje veľa výnimiek z pravidiel a mnohé ešte neznáme. Avšak všetci chovatelia psov by sa mali pokúsiť porozumieť najjednoduchším v genetickej teórii Mendela - "dominantný" alebo "recesívny" gén alebo vlastnosť. To je ľahké pochopiť a môže byť aplikované priamo na chov psov, pretože značný počet závažných nedostatkov plemena je spôsobený recesívnymi génmi.
Recesívny gén sa nemusí prejavovať v jednej alebo viacerých generáciách, kým sa nájdu dva identické recesívne gény z každého rodiča. Psy, ktoré majú len jeden recesívny gén, túto vlastnosť neukazujú, pretože manifestácia recesívneho génu bude maskovaná vplyvom dominantného génu spárovaného s ním. Takéto psy / nosiče recesívneho génu / môžu byť pre toto plemeno nebezpečné, ak tento gén určuje výskyt nežiaducich znakov tým, že ich prenesie na svojich potomkov a zostane v pleme. Ak ste náhodou združili dvoch psov, nositeľov takéhoto génu, dajú časť potomkov nežiadúcim znakom.
Dominantný gén sa vždy jasne a zvonka prejavuje zodpovedajúcim znakom. Preto dominantné gény, ktoré nesú nežiaduce znaky, sú pre chovateľa omnoho nižšie riziko, pretože ich prítomnosť sa vždy prejavuje, aj keď dominantný gén funguje bez partnera.
Ale zrejme, aby sa všetko komplikovalo, nie všetky gény sú absolútne dominantné alebo absolútne recesívne. Niektoré gény sú dominantnejšie ako iné a naopak.
Párovanie nie vždy poskytuje pomer v presnom súlade s očakávanými priemernými výsledkami a na získanie spoľahlivého výsledku z tohto párovania je potrebné vyrobiť veľké vrhy alebo veľké množstvo potomkov v niekoľkých vrhoch.
Genetická tabuľka znázorňujúca jednoduchú interakciu dominantných a recesívnych charakteristík podľa Mendela
Legenda: A - dominantný jednorodičový gén
a - recesívny gén jedného rodiča
AA - dvojica dominantných génov, jeden od každého rodiča
aa - pár recesívnych génov, jeden od každého rodiča
Aa je dominantný gén jedného rodiča a recesívny druh, ktorý tvorí dominantný recesívny pár génov
Vysvetlenie možností:
1. Dvaja rodičia majú dva dominantné gény, takže všetci ich potomkovia budú týmto znakom "čistí" - AA
2. Jeden rodič je "čistý" dominantným faktorom / AA /, druhá len vyzerá "čistá", ale nesie recesívny gén (Aa). Všetci potomkovia budú vyzerať čisto z recesívneho znaku, ale v skutočnosti polovica potomkov bude naozaj čistá, má dva dominantné gény (AA), druhá polovica bude "čistá", ale každá z nich bude mať recesívny gén (AA / a bude jeho nosičom. Pomer AA: AA - 1: 1
3. Jeden rodič prejaví sa zvonka dominantným znakom, ale je nosičom recesívneho génu (Aa). Pri párení s partnerom, ktorý nesie dva recesívne gény a prirodzene prejaví tento rys, polovica potomkov bude podobná prvému partnerovi / Aa /, druhá polovica druhého / aa /. Takže celé vrh bude nosičom tohto génu, iba prvý polčas - skrytý, druhý explicitný.
4. Obaja rodičia prejavujú dominantný znak, ale každý z nich má recesívny gén, ktorý sa nezjavuje. Vrh bude jedným z potomkov čisto "dominantné", ktoré majú dve A / AA /, dvoch potomkov, zrejme ukazuje dominantný vlastnosť a to nie je odlišný od toho prvého, skrytý bude niesť recesívne zvláštnosť, s Aa, a jeden z potomkov bude recesívny znamenie, že má dva recesívne gény aa. To znamená, že pomer potomstva vykazujúcich znak definovaný génu je 3: 1, a pravú definíciu nosiče recesívne a dominantné Hell budú nasledovne: AA: Aa: aa - 1: 2: 1
5. V tomto páre je jeden rodič "čistý" dominantný / AA /, druhý má dva recesívne gény / aa /. Všetky vrhy budú vyzerať ako dominantný rodič, ale každý z potomkov bude mať recesívny gén (Aa /
6. Obaja rodičia vykazujú recesívny príznak, pretože nesú dva recesívne gény / aa /. Všetky vrhy tohto páru budú mať tento znak v explicitnej forme a odovzdávať ho svojim potomkom, kým nebudú spárované s partnerom s dominantným génom. V tomto prípade bude výsledok párovania rovnaký ako v páre 3 a 5.
Zdroj: kniha Hillery Harmar "Psy a ich chov"
Aby sme si hrubo predstavili, čo potomkovia môžu pochádzať od daného páru výrobcov, je potrebné poznať základné pravidlá dedičnosti a ktoré vlastnosti sú recesívne a ktoré sú dominantné.
Tu je zoznam najbežnejších znakov identifikovaných recesívnymi génmi pre väčšinu plemien:
Snack, prekrytie, ľahký nos, svetlé oči, dĺžka končatiny, krátka papuľa, malé uši, vztýčené uši. Mnohé z týchto závažných vrodených chýb sú tiež riadené recesívne gény: rázštep pery, rázštep podnebia, zapletenej chvostom, hluchota, vrodené prietrže, vlčie pazúriky, strach z ostrých zvukov, močenie, keď vzrušený, Entropium storočia, ektropium, albinizmus, šedý zákal, atď.
Tvar lebky a hlavy sa zdedil podľa princípu Nielson-Ele. Jednoznačne pôsobiace gény sa nazývali polymérne a samotné polymérové dedičstvo / Široká horná čeľusť dominuje úzkym, šírka základne lebky je tiež zdedená. Široká forma lebky a dolnej čeľuste dominuje úzkemu. nie úplné nadvlády má obdĺžnikovú hlavu nad klinovým tvarom - a úplná dominancia má hlavu v tvare buldoka nad normálnym.
Výška a hmotnosť sa zdedia medzičasom.
HH visiace uši dominujú stojace hh, ale existujú aj stredné možnosti - Hh semi-visiace uši
Prítomnosť pazúrikov na zadných končatinách dominuje ich absencii.
Narrowwness dominuje nad šírkou.
Taxonog dominuje normálnej štruktúre nohy.
Extrémne možnosti - ľahké a ťažké kosti dominujú medziľahlé typy.
Hladkosrstý je dominantný vo vzťahu k dlhosrstým.
Zdobenia chĺpkov sú dominantné vo vzťahu k ich absencii.
Zvrat vlasov je dominantný vzhľadom na ich neprítomnosť.
Kniha Hillaryho Harmara "Psy a ich chov"
Aby sme pochopili význam chovu, musíme začať od samého začiatku - od narodenia nového života. Bunka je jednotkou života a pochádza z inej žijúcej bunky. Každé zviera je výsledkom zlúčenia dvoch rodičovských buniek v jednej dcére.
Keď sa narodí nový život, jedna a jedna spermiová bunka (bunka oka) preniká cez vaječný škrup (materská bunka). Plášť vajíčka potom okamžite zahustí a zabraňuje prenikaniu iných spermií. Ale pred zrenie spermatozómu sa mužská bunka obsahujúca dvojitú sadu chromozómov rozdelí na dve, takže polovica chromozómov (jedna sada) prechádza do jednej časti a druhá polovica (rovnaká sada) do druhej. Preto v každom pohlavnom pohlade samčí bunka-spermie obsahuje jednu sadu chromozómov. Ženská reprodukčná bunka, vajíčko, sa tvorí podobným spôsobom a obsahuje rovnaký súbor chromozómov. Kombinácia dvoch polovíc - obidvoch zárodočných buniek - spermie a vajíčka - v ktorých každý chromozóm nájde "partnera" z inej bunky, tvorí novú bunku, z ktorej sa môže vyvinúť nový živý tvor, nesúci chromozómy oboch rodičov, opäť dvojitý súbor chromozómov.
Ale nie všetky chromozómy nájdu svojich "partnerov" vo fúzii rodičovských zárodočných buniek, napríklad v chromozómoch, ktoré určujú pohlavie.
Ženská bunka má iba jeden typ pohlavného chromozómu, ktorý sa nazýva chromozóm X. Samčia zárodočná bunka môže mať X a Y chromozómy, ale nie obe. Ktorý z nich sa spojí so ženským X chromozómom je náhodou. Ak je to X chromozóm, potom bude zárodok samica "XX". Ak ide o Y-chromozóm, embryo bude samčí "XY", čo je charakteristické pre všetky cicavce.
V momente spojenia dvoch polovíc rodičovských buniek začína nový život. V takej malej klietke je prototyp budúceho psa, z neho sa vytvára krv, nervový systém, vlna, svaly, kosti, farba očí závisí od nej, podlaha - v skutočnosti celý budúci pes.
Prvá bunka je rozdelená na dve, potom na štyri, osem, šestnásť atď. A naďalej sa rýchlo množia. Každá bunka má jadro, ktoré je najdôležitejšou zložkou. Obsahuje sieť vláknitých štruktúr, ktoré sa tiahnu do vlákien nazývaných chromozómy. Páry chromozómov (jeden z každého rodiča) sa držia spolu a nesú veľa ultramikroskopických jednotiek nazývaných gény, sú tiež spárované.
Gén je jednotka dedičnosti a jediný fyzický článok v reťazci, ktorý spája generácie. Gény sú rozdelené takým spôsobom, že každý z nich má osobitné miesto na vlastnom chromozóme. Každý gén nesie svoje dedičné vlastnosti a riadi manifestáciu rôznych fyzických a mentálnych charakteristík v dospelom živote. Pes má 78 chromozómov (39 párov), ktoré obsahujú tisíce génov. Gény zostávajú nezmenené z generácie na generáciu, pokiaľ nedôjde k mutácii. Pri chromozomálnej dedičnosti je vplyv rodičov rovnaký, s výnimkou prípadov dedičnosti spojených s pohlavím. Gény jedného chromozómu sú spravidla zdedené spoločne. Každá špecifická vlastnosť alebo vlastnosť živého organizmu je určená jeho genómom, z ktorého polovica je zdedená jedným z rodičov, polovica druhého. Pre rozvedeného je najdôležitejšie vedieť o tých nosných génoch pre postavy, ktoré majú dominantný recesívny vzťah.
Recesívne gény
Recesívny gén (tj vlastnosť, ktorú určuje) nemôže spraviť jednu alebo viacero generácií, kým sa nedosiahnu dva identické recesívne gény od každého z rodičov (náhle prejavy takéhoto znaku u potomkov by sa nemali zamieňať s mutáciou).
Psy, ktoré majú len jediný RECESSIVE GENE - determinant akéhokoľvek znaku - neukazujú túto vlastnosť, pretože účinok recesívneho génu bude maskovaný prejavom vplyvu páru DOMINANT GENE. Takéto psy (nosiče recesívneho génu) môžu byť pre toto plemeno nebezpečné, ak tento gén určuje vzhľad nežiaducich znakov, pretože budú prenesené na jeho vlastný kameň a tieto budú pokračovať; a gén sa takto zachováva v chove. Ak sa náhodou alebo bezmyšlienkovo spája Dva nosiče takého génu, dajú časť potomkov nežiadúce vlastnosti.
Genetická tabuľka znázorňujúca jednoduchú interakciu
dominantných a recesívnych charakteristík spoločnosti Mendel
Možnosť párenia .... Rodičia ............... Potomkovia
1 ............. ......................... AA x AA ....... ..... ..... ... AA AA AA AA
2 ...... .......... ...... ... ............. AA x Aa ....... ..... .... AA AAA Aa Aa
3 ......... .......... ...... ... .......... Aa x aa ...... ...... ... .aaaaaaaa
4 ......... .......... ......... ... ....... Aa x Aa ... ... ..... ... AA Aa Aa aa
5 ............ .......... ...... ... ...... .... AA x aa ...... ..... ... .aaaaaaaa
6 ............ .......... ...... ... ... ... .... aa x aa ...... ...... ... .aaaaaaaa
Dominantný gén od jedného rodiča
recesívny gén od jedného rodiča
AAA - dvojica dominantných génov, jeden od každého rodiča
aa - pár recesívnych génov, jeden od každého rodiča
Aaa je dominantný gén z jedného rodiča a recesívny gén od druhého,
zložiek dominantného recesívneho páru génov
Vysvetlenie možností:
1. Obaja rodičia majú dva dominantné gény, takže všetci ich potomci budú na tomto základe "čistí" - všetci AA.
2. Jeden rodič je "čistý" dominantným faktorom (AA), druhý vyzerá "čistý", ale nesie recesívny gén (AA). Z tohto dôvodu budú všetky potomkovia vyzerať "čisté" z recesívne znak, ale v skutočnosti je polovica detí skutočne "čisté", s dvoma dominantnými gény (AA), druhá polovica bude vyzerať "čisté", ale každý z nich bude mať recesívne gén, t.j. bude jeho nosičom (Aa). Pomer AA: Aa - 1: 1
3. Jeden rodič má zvonka dominantnú vlastnosť, ale je nosičom recesívneho génu (Aa). Pri párení s partnerom, ktorý nesie dva recesívne gény a prirodzene externe ukazuje túto vlastnosť, polovica potomkov bude podobná prvému partnerovi (Aa), druhá polovica druhému. Takže všetky vrhy budú nositeľom tohto znaku, iba prvé budú skryté nosiče a druhé - explicitné. To je obzvlášť dôležité na pochopenie pre tie prípady, keď je znak určený recesívnym génom zvlášť nežiadúcim pre plemeno.
4. Obaja rodičia vykazujú dominantnú vlastnosť, ale každý nesie recesívny, neexistenčný gén. V vrhu bude jedna časť potomkov čisto "dominantnej", ktorá má dve A (AA); dve časti potomkov, ktoré navyše vykazujú dominantnú vlastnosť a teda sa nelíšia od prvej, budú mať aj recesívny znak, ktorý má Aa; jedna časť potomkov vykazuje recesívny znak, ktorý má dva recesívne gény (aa). To znamená, že pomer potomstva vykazujúcich znak definovaný génu bude 3: 1, a pravý rozloženie nosičov recesívnych a dominantných génov by bolo nasledovné: AA: A: A - 1: 2: 1.
5. V tomto páre - jeden rodič je "čistý" dominantný (AA), druhý má dva recesívne gény (aa). Všetok vrh bude vyzerať ako dominantný rodič, ale každý z potomkov bude nositeľom recesívneho génu (Aa), a teda jeho znakom určeným.
6. Obaja rodičia vykazujú recesívny znak, pretože každý z nich má dva recesívne gény (aa). Všetky vrhy tohto páru tiež budú mať tento znak a odovzdávať ho potomkom explicitne, kým nebudú spárované s partnerom nesúcim dominantný gén. V tomto prípade bude výsledok párovania rovnaký ako v páre 3 a 5.
Očakávaný pomer štiepenia potomstva na konkrétnom základe je približne odôvodnený vrhom najmenej 16 šteniat. Pri vrhu bežnej veľkosti - 6-8 šteniat - možno hovoriť len o väčšej alebo menšej pravdepodobnosti prejavu znaku určeného recesívnym génom pre potomkov určitého páru výrobcov s známy genotyp.
Dominantné gény
Prítomnosť dominantného génu sa vždy jasne a zvonka prejavuje zodpovedajúcim znakom. Preto dominantné gény, ktoré nesú nežiaduci znak, predstavujú pre chovateľa oveľa nižšie riziko ako recesívne, pretože ich prítomnosť sa vždy prejavuje, aj keď dominantný gén "funguje" bez partnera (t.j. Aa).
Ale zrejme, aby sa skomplikovali záležitosti, nie všetky gény sú absolútne dominantné alebo recesívne. Inými slovami, niektoré sú dominantnejšie než iné a naopak. Napríklad niektoré faktory, ktoré určujú farbu srsti, môžu byť dominantné, ale stále sa neobjavujú smerom von, ak nie sú podporované inými génmi, niekedy dokonca recesívnymi.
Párovanie nie vždy poskytuje pomer v presnom súlade s očakávanými priemernými výsledkami a aby sa získal spoľahlivý výsledok z tohto párovania, je potrebné urobiť veľké vrhy alebo veľké množstvo potomkov v niekoľkých vrhoch.
Niektoré vonkajšie znaky môžu byť "dominantné" v niektorých horách a "recesívne" v iných. Ďalšie črty môžu byť spôsobené viacerými génmi alebo pologénmi, ktoré nie sú len dominantnými faktormi alebo Mendelov recesívnymi. V dôsledku toho sa genetika stane príliš zložitou, aby ju chápali priemerní chovatelia psov!
mutácie
Mutácia - náhla zmena génu. To sa prejavuje v prvej generácii potomkov, ak je dominantný mutantný gén. Ale recesívny mutantný gén môže byť tajne zdedený niekoľko generácií, kým dvaja nosiči takého génu nie sú vybraní v rodičovskom páre. Iba potom sa objaví potomok, ukazujúci výsledok mutácie tohto génu.
Mnoho vonkajších zmien je spôsobených mutáciami. Klasickým príkladom toho sú plemená so štvorcovými tvárami, ako napríklad predčasní mastiffi pred stovkami rokov, a všetky plemená so skrátenou tvárou, ako sú Pekingese, mopslíky, buldogy. Plemená ako basy, pekingi a jazvečíci trpia hereditárne fixovanou mutáciou, ktorá spôsobuje deformitu známu ako achondroplázia (abnormálny vývoj tubulárnych kostí končatín pred narodením, vyjadrený poklesom ich dĺžky).
Mutácie sú prirodzené, ale môžu byť tiež spôsobené umelo, napríklad ionizujúcim žiarením (žiarením). Lieky a jedy môžu byť ďalšou príčinou a zvyčajne spôsobujú škodlivé mutácie. Vplyv prostredia môže tiež ovplyvniť frekvenciu mutácií. Zaujímavé je, že mutácie sú zdedené, to znamená, že sú vždy reprodukované, takže nové charakteristiky alebo znaky sa môžu objaviť neustále.
Smrteľné gény
Toto sú gény, ktoré spôsobujú smrť organizmu skôr, než dosiahne pubertu. Smrteľné gény sú recesívne, tu sú niektoré príklady prejavu ich vplyvu: "rozštiepenie pery a rozštiepeného podnebia" - chyba vo vývoji hornej čeľuste, hemofília - nedostatok krvnej schopnosti koagulovať, "vstrebávanie plodov" vo vonkajšej prosperujúcej fenke atď.
Semilatálne gény, napríklad gény, ktoré určujú obojstranný kryptorchidizmus, sa nakoniec stanú smrteľnými pre plemeno v dôsledku jeho vyhynutia. Šteňatá s "vlčím ústom", ak nie sú v prevádzke, nemôžu nasávať a preto zomierajú. Farba šedo-modrej farby s čiernymi škvrnami je spojená s polo-letálnym génom a ak je zdedený potomkom oboch rodičov, potom tento potomok môže byť slepý, hluchý alebo neplodný. Z tohto dôvodu sa dva psi tejto farby nikdy nespávali. Prakticky by bolo najlepšie považovať túto farbu za diskvalifikujúce vo všetkých plemenách.
Rozumné chovanie psov sa vykonáva kombináciou dvoch hlavných typov chovu - INBRIDING a LYNBRIDING, ale z času na čas je potrebný primeraný AUTOCROSSING. V priemere bude potomstvo vo všetkých druhoch chovu takmer rovnaké, ale kde sa použilo inbreeding, najmä v nadmernom rozsahu, potomstvo bude buď výrazne lepšie, alebo oveľa horšie.
Malo by sa tiež pamätať na to, že ani najvýznamnejší chovný pes nebude v genetickom zmysle rovnako vhodný pre všetky feny.
inbreeding
Inbreeding je spárovanie blízkych príbuzných, napríklad otec s dcérou, matka so synom, brat s sestrou atď. Inbreeding by sa nikdy nemalo používať, ak neexistuje istota, či sú hospodárske zvieratá absolútne zdravé, fyzicky aj duševne. Týmto spôsobom chovu je prísne povinné odmietnutie všetkých plemenných materiálov, ktoré nespĺňajú normy. Ak najhoršie šteňatá nie sú zničené, nemali by vyrábať rodokmeň a samozrejme by sa z nich nikdy nemali dostať potomkovia.
Niektoré z najlepších hospodárskych zvierat na svete koní, kráv, ošípaných, psov a iných domácich zvierat boli získané inbreeding. Ale použitie inbreeding bez dostatočných znalostí je možné zničiť plemeno už niekoľko generácií. Možno, v jednej generácii a samozrejme v dvoch inbreeding, ukáže všetky chyby, ktoré sú geneticky začlenené do plemena. Malo by sa opäť zdôrazniť, že eliminácia všetkých neúspešných potomkov z ďalšej plemenárskej práce je veľmi dôležitá počas vtáctva.
Noví chovatelia psov, ktorí nepoznajú psov uvedených v rodokmeni, aspoň pred štyrmi generáciami, je nevhodné pokúsiť sa použiť inbreeding. Rizikové experimenty sú veľmi drahé a môžu priniesť veľké sklamanie. Stáva sa, že inbreeding odhaľuje nedostatky predkov oddelených viacerými generáciami.
linebreeding
Linegenie je ako inbreeding - príbuzní sú tiež spolu, ale ich spoločný predchodca sa nachádza len v tretej - štvrtej generácii. Samozrejme, čím vzdialenejší je vzťah, tým viac sa bude meniť druh prijímaných potomkov. Preto je pre priemerného rozvedenca rozumnejšie chrániť seba a používať tesné linebreeding pre svojich psov. S línie, ako aj s inbreeding je veľmi dôležité odstrániť zo šľachtenia všetky psy, ktoré nespĺňajú štandard.
kríženie iných rás
Prekročenie je párovanie nezávislých výrobcov. Použije sa, keď je potrebné zaviesť niektoré špecifické charakteristiky z inej línie alebo opraviť nedostatok, ktorý prenikol do línie v častiach fyzických alebo duševných charakteristík.
Niekedy veľmi prepracované párovanie prináša neuspokojivé plemená a nakoniec sa zapletený naplno očakáva od nejakej zmesi šteniatok, ale náhle sa pes a fena prekvapujúco vyhovujú navzájom a dávajú vynikajúce krásne šteniatka. Tak, jeden kompletný ďalšiemu kríženie môže napraviť vadu, ale pozor - potomkovia takých krížov sú zriedka prominentnú psoch a suky a nerobia zo seba veľké potomstvo, pokiaľ neboli spárované "späť", tj lineárne inbridirovany samca psa či suku .. ich vlastnú krvnú líniu, ktorá má dominantné gény pre požadované vlastnosti.
Ale jednoduché kopírovanie úspešných chovateľských programov neprinesie chovateľovi očakávaný úspech. Niekedy rozvod nevedomosti o kvalite predkov svojich chovných psov prináša plemenu toľko škôd, že v čase, keď si uvedomí rozsah poškodenia, je často príliš neskoro na to urobiť čokoľvek. Preto je chovateľ psov pred vykonávaním chovu psov povinný čo najlepšie sa zoznámiť s vybraným plemenom, o najbežnejších nedostatkoch, o najlepších a najhorších predkoch chovných psov, s ktorými sa bude chovať.
Najčastejšou príčinou nedostatkov a odchýlok od normy sú recesívne gény. S približnou presnosťou je možné zvážiť:
ak potomok vykazuje charakteristiku, ktorú žiadny z rodičov nemal,
táto vlastnosť je určená recesívnym génom
Napríklad, ak obaja rodičia majú tmavé oči alebo špičky nosa a šteňa má svetlo, potom obaja rodičia mali predkov s recesívnym génom, ktorý určuje takýto znak, a oni zdedili tento gén. Pri párovaní týchto psov dohromady sa gény spojili - v dôsledku toho rodičia s tmavými očami poskytli ľahký pohľad na potomstvo. Ak je toto potomstvo prepojené s podobným psom so svetlými očami, s rodičmi s tmavými očami, žiadne z ich šteniatok nebude mať tmavé oči, Tento princíp platí aj pre defekty, ktoré sú definované jednoduchým recesívnym génom, ako je rozštiepenie podnebia alebo hluchota. Z toho je zrejmé, aký nedostatok, určený recesívnym génom, je ľahko zachovaný a rozšírený v pleme, najmä ak je nosičom populárny chovný pes, ktorý môže vo svojom živote produkovať niekoľko stoviek potomkov. Dokonca aj keď má iba jeden recesívny gén, ktorý identifikuje vážnu chybu, je jasné, že bez toho, aby túto chybu dokázal, môže rýchlo rozšíriť túto chybu v plemenách. V čase, keď sa objaví, nebude možné nič opraviť, zvlášť ak sa potomky tohto psa široko používajú.
Keď chovné psy majú tendenciu k extrémnym charakteristikám, plemeno sa takmer určite zhoršuje. Psy z tých plemien, ktoré napriek ľudskému zásahu zostávajú prirodzené svojim divokým predkom, netrpia veľa z rôznych chýb fixovaných v iných pleciach charakteristickú vlastnosť, Ak chovatelia psov boli vedomí utrpenia a utrpenia, ktoré spôsobili svojim psom, nehovoriac o tom, že platia veterinárne služby na opravu charakteristík exotických plemien, potom by sa už rozhodli, že by bolo múdrejšie meniť normy. Chovatelia psov ľahko prestávajú myslieť, pokiaľ ide o svojich psov, aj keď sú s nimi veľkodušní, ale rýchlo sa stávajú slepými a často prehliadajú tragédie, ktoré chovajú. Môžeme uviesť len niekoľko príkladov, úplný zoznam by bol príliš dlhý:
Vždy sú ťažké pôrody, keď lebky a panvové otvory nezodpovedajú svojim rozmerom (napríklad u bostonských teriérov a francúzskych buldokov);
- skrátené končatiny zmenia polohu lalokových kostí panvy, zatiaľ čo panvový otvor je príliš nízky a ak majú psy tohto plemena lakované brucho, porodenie bude ťažké (napríklad škótsky teriér);
- príliš dlhý chrbát a bedrá poskytujú dodatočné zaťaženie chrbtice, čo sa odráža v ochorení medzistavcovej chrupavky (jazvečíkov, basiek);
- príliš krátke čapíky spôsobujú ťažkosti s dýchaním, tieto plemená veľmi často, aspoň častejšie ako iné, spôsobujú narodenie šteniatok s ústami vlkov (Pekingese);
- prehnané voľné kože tvoria hlboké záhyby, kde sa často vyskytuje vyrážka pri plienkach, viečka pokryté pokožkou prestávajú chrániť oči, kedy dochádza k pretrvávajúcej chronickej konjunktivitíde;
- príliš úzky zvukový kanál a vlna, ktoré tu rastú, vytvárajú podmienky pre pretrvávajúce ochorenia uší (fox teriérov, pudlíky) ... a mnoho ďalších ...
Bohužiaľ, príroda nesmie hrať svoju správnu úlohu pri chove psov, všetky slabé šteniatka sú zachránené, kŕmené, a potom získajú potomky z nich, zatiaľ čo v prírode budú musieť zomrieť. Zákon prírody "prežije najschopnejší k životu" nemôže byť beztrestne porušený. Je dobré, že mnoho psov s dedičnými poruchami je neplodné alebo menej plodné, takže v tomto pleme nepretrvávajú tie najstrašnejšie nedostatky, ako to mohli s pomocou "dobrých" psích chovateľov.
U psov, ako aj u iných zvierat je vždy správne spájať to najlepšie s najlepším, až potom môže šľachtiteľ dúfať, že dosiahne najlepší výsledok. Existuje veľa charakteristík a charakteristík, ktoré nie sú určené jediným párom génov, ale veľkým počtom génov. Napríklad v chrtach nie je možné predvídať rýchlosť jazdy, pretože je určená kombináciou veľkého počtu dedičných faktorov, to znamená, že neexistujú žiadne dominantné alebo recesívne príznaky pre rýchlosť jazdy. Je však známe, že v niektorých rodinách alebo rýchlostiach je rýchlosť vyššia, takže opäť spárovanie najlepších s najlepšími na dlhú dobu určite dáva to najlepšie.
Pozorovaním chovateľa sa čoskoro zistí, ktoré gény sú dominantné. Podľa môjho názoru je však oveľa dôležitejšie, aby majiteľ psa vedel, ktoré gény sú recesívne, pretože vyžadujú najväčšiu pozornosť pri chove. Bohužiaľ nie všetky rysy sú definované rovnako pre všetky plemená dominantnými alebo recesívnymi génmi. Platí to najmä pre farby srsti.
Možno sa opäť zopakuje, že ak šteniatko má znaky, ktoré nie sú vlastné žiadnemu z jeho rodičov, potom tieto znaky sú kontrolované recesívnymi génmi.
Tu je užitočné poskytnúť zoznam najbežnejších charakteristík určených recesívnymi génmi pre väčšinu plemien:
Nosh
majúce vystupujúcou čeľusť
Ľahký nos
Svetlé oči
Dĺžka končatiny (pre väčšinu plemien)
Krátka papuľa
Malé uši
Vztýčené uši (pre väčšinu plemien)
Dlhá vrstva (recesívna pre hladkú vrstvu)
Hladká (krátka) vlna (recesívna pre dlhosrstú)
Mäkká vlna (recesívna pre tvrdé vlasy)
Rovná vlna (recesívna pre kučeravé)
Mnohé z vážnych dedičných chýb sú tiež kontrolované génmi, ktoré sú recesívne pre väčšinu plemien, napríklad:
rázštep pery, rázštep podnebia, zálomok, hluchota, vrodená prietrž, vlčie pazúriky, strach z ostrých zvukov, močenie, keď vzrušený, Entropium storočia, ektropium, albinizmus, ktoré majú sklon tvoriť kamene v močovom mechúre, hemofíliu, zahustenie a zabalenie pery, šedý zákal a pr.
Farba vlny a pigmentácia pre mnohé plemená sú pevne stanovené požiadavkami normy. Tieto znaky sú jedným z najťažších z hľadiska dedičnosti, pretože kontrolné gény môžu byť dominantné pre jedno plemeno a recesívne pre iné, zatiaľ čo kontrolujú rovnakú farbu. Pokiaľ ide o pigmentáciu, všetky plemená majú hnedý alebo svetlý nos, jasné oči, všeobecné oslabenie pigmentácie je určené recesívnymi génmi.
Ak je chov potrebný na vedenie farby, musíte vytvoriť zoznam všetkých recesívnych farieb známych v tomto pleme. Pre tých, ktorí majú skutočný záujem o chov podľa farby, dôrazne odporúčam knihu Clarence Litt "Dedičstvo vlnových farieb u psov".
zovšeobecnenie
Jednou z najväčších tragédií, ktorá chápe majiteľa psa, je takzvaná "škôlka slepoty". Tento termín znamená, že chovateľ psov nevidí nedostatky psov chovaných, ale je veľmi dobrý, keď vidí nedostatky hospodárskych zvierat iných chovných staníc. Existuje však aj ďalšia, možno aj väčšia vada chovných psov, tzv. "Chovnej slepoty". K tomu dochádza vtedy, keď nedostatok nielen prechádza, ale je už v ňom zakorenený a odborníci v kruhu to nechali prechádzať, pretože je len veľmi málo psov, ktoré nemajú taký nedostatok. Nakoniec sa táto nevýhoda stane prijateľnou pre plemeno. Tak to bolo napríklad s dislokáciou kolenného kĺbu.
Úprimnosť psa chovateľa pri rozpoznávaní chýb v otvorenej diskusii je veľmi dôležitá, ale hlavnou zodpovednosťou za chov najlepších psov je v skutočnosti odborníci na výstavy. Ak nikdy nezaťažia prvého psa vážne nevýhodou, chovatelia psa, samozrejme, nebudú utrácať peniaze a vystavovať takýchto psov, pretože pochopia, že pes s týmito nedostatkami nemá šancu na prehliadku. Ale kým chovatelia psov môžu zvíťaziť s nezdravými psami, tak s ohľadom na ľudskú prirodzenosť budú takí psi nielen vystavení, ale budú sa používať aj na chov. Niektorí odborníci s potešením píšu vo svojich správach, že daný pes, alebo dokonca všetci psi určitého plemena, ktoré boli vyšetrení, sú pozoruhodne zdravé, zatiaľ čo fyzicky väčšina z nich bola mimoriadne nezdravá.
Je to medvedia plemena, ale rovnako tak lichotivé spôsob, ako na šnupavý tabak vážny chovateľov, ktorí striktne dodržiavať, aby zabezpečili, že hornina nie je "priznať" nedostatky, a to najmä tých, ktorí porušujú zdraví a hrozí, že psie existencie, rovnako ako sivý zákal, Entropium storočia súmraku slepoty a dysplázie kĺbov.
Existuje príliš veľa psov a možno aj menej a lepšie, ale bohužiaľ "pesovka" bola vždy pre politikov z psího chovu celkom atraktívna. Avšak v priebehu času sa napriek dôležitým odborníkom a nevedomosti chovateľov psov väčšina plemien zlepšila a naďalej zlepšovala, najmä vzhľad, Takýto záver možno urobiť porovnaním dnešných predstaviteľov plemien s fotografiami prijatými pred 50 rokmi.
Bohužiaľ, existuje extrémne veľa psích chovateľov, ktorí "nevidia" psa, a preto sú úplne úprimne neschopní si všimnúť nedostatky a následne ich zásluhy. Osoba sa naozaj narodila s prirodzenou schopnosťou vidieť zviera, potom si môže vybrať dobrého psa v každom chove. Odborníci sa taktiež narodia, nie sú robení, aj keď sa samozrejme môžu hromadiť znalosti.
Najťažšie určiť najlepšieho psa, keď si musíte vybrať z veľkého počtu zlých alebo stredných psov. Vo všetkých ohľadoch je nádherný exemplár s jednou viditeľnou chybou lepší ako priemerný pes, ktorý má veľmi dobré a veľmi zlé vlastnosti. Je zrejmé, že pri ďalšom šľachtení je jednoduchšie použitie potomkov týchto psov sa zbaví jednej nevýhody skôr než snažiť sa zaviesť do plemena súčasne veľa dobrých vlastností.
Recesívne gény
Recesívneho génu (tj znamenie, ktoré je definované) nesmie prejaviť v jednom alebo mnoho generácií, kým spĺňajú dve identické recesívny gén od každého z rodičov (náhly prejav tohto znaku u potomstva by sa nemala zamieňať s mutáciou).
Psy, ktoré majú len jeden RECESSIVE GENE - determinant akéhokoľvek znaku - nebudú vykazovať túto vlastnosť, pretože účinok recesívneho génu bude maskovaný prejavom vplyvu DOMINANTOVÉHO GENE párovaného s ním. Takéto psy (nosiče recesívneho génu) môžu byť pre plemeno nebezpečné, ak tento gén určuje výskyt nežiaducich znakov, pretože sa bude prenášať do svojho pôvodu a tým ďalej, a preto zostane v pleme. Ak sa náhodou alebo bezmyšlienkovo spája Dva nosiče takého génu, dajú časť potomkov nežiadúce vlastnosti.
Genetická tabuľka znázorňujúca jednoduchú interakciu dominantných a recesívnych charakteristík podľa Mendela
Legenda:
Dominantný gén od jedného rodiča
A -? - recesívny gén od jedného rodiča
AA - dvojica dominantných génov, jeden od každého rodiča
aa -? ? - pár recesívnych génov, jeden po druhom od rodiča
Ah -? - dominantný gén od jedného rodiča a recesívny gén od druhého, tvoriaci dominantný recesívny pár génov.
Vysvetlenie možností:
1. Obaja rodičia majú dva dominantné gény, takže všetci ich potomci budú na tomto základe "čistí" - všetci sú AA.
2. Jeden rodič je "čistý" v dominantnom faktore (AA), druhý vyzerá "čistý", ale nesie recesívny gén (Aa). Všetci potomkovia budú vyzerať čisto z recesívneho znaku, ale v skutočnosti polovica potomkov bude naozaj "čistá", má dva dominantné gény (AA), druhá polovica bude vyzerať čisto, ale každý z nich bude mať recesívny gén, tj bude jeho nosičom (Aa). Pomer AA: Aa - 1: 1.
3. Jeden rodič má zvonka dominantnú vlastnosť, ale je nosičom recesívneho génu (Aa). Pri párení s partnerom, ktorý nesie dva recesívne gény a prirodzene externe ukazuje túto vlastnosť, polovica potomkov bude podobná prvému partnerovi (Aa), druhému druhému druhému. Takže všetky vrhy budú nositeľom tohto znaku, iba prvé budú skryté nosiče a druhé - explicitné. To je obzvlášť dôležité na pochopenie pre tie prípady, keď je znak určený recesívnym génom zvlášť nežiadúcim pre plemeno.
4. Obaja rodičia vykazujú dominantnú vlastnosť, ale každý nesie recesívny, neexistenčný gén. V vrhu bude jedna časť potomkov čisto "dominantnej", ktorá má dve A (AA); dve časti potomkov, ktoré navyše vykazujú dominantnú črtu, a teda sa nelíšia od prvej, budú niesť recesívny znak, ktorý má Aa; jedna časť potomkov vykazuje recesívny znak, ktorý má dva recesívne gény (aa). To znamená, že pomer potomkov vykazujúcich znak určený genómom A bude 3: 1 a skutočná distribúcia nosičov recesívnych a dominantných génov bude nasledovná: AA: Ah: aa - 1: 2: 1.
5. V tomto páre - jeden rodič je "čistý" dominantný (AA), druhý má dva recesívne gény (aa). Celý vrh tohto páru bude tiež nosiť tento rys a odovzdávať ho ďalej potomkom v explicitnej forme, kým nebudú spárované s partnerom nesúcim dominantný gén. V tomto prípade bude výsledok párovania rovnaký ako v páre 3 a 5.
6. Obaja rodičia vykazujú recesívny znak, pretože každý z nich má dva recesívne gény (aa). Celý vrh tohto páru bude tiež nosiť tento rys a odovzdávať ho ďalej potomkom v explicitnej forme, kým nebudú spárované s partnerom nesúcim dominantný gén. V tomto prípade bude výsledok párovania rovnaký ako v páre 3 a 5.
Očakávaný pomer štiepenia potomstva na konkrétnom základe je približne odôvodnený vrhom najmenej 16 šteniat. V prípade vrhu bežnej veľkosti - 6-8 šteniat - možno hovoriť len o väčšej alebo menšej pravdepodobnosti prejavu znaku určeného recesívnym génom pre potomkov určitého páru výrobcov so známym genotypom.
Z knihy Tajomstvo ľudského dedičstva autora Afonkin Sergey YuryevichDominantné a recesívne gény Predstavte si dva homológne chromozómy. Jedna z nich je materská, druhá otcovská. Kópie génov nachádzajúcich sa na rovnakých segmentoch DNA takýchto chromozómov sa nazývajú alelické alebo jednoducho alely (grécky alios je iný). Tieto kópie
Z knihy Tajomstvo ľudského dedičstva autora Afonkin Sergey YuryevichGény starnutia veku sú nepríjemné a každý rok sa zhoršuje. Diane Cooper Ľudia nechcú žiť navždy. Ľudia jednoducho nechcú zomierať. Stanislav Lem - Nesmrteľné bunky existujú - Programovaná smrť - Hodiny starnutia - Choroby
Z knihy Evolution of Man. Kniha 2. Opice, neuróny a duša autoraGény a správanie Fakty získané neurobiológmi hovoria o materiálnej, neurologickej povahe psychiky. Ale aby sa duša vo všetkých jej prejavoch mohla vyvíjať, toto, striktne povedané, stále nestačí. Nie všetky znaky sa môžu vyvíjať, ale iba
Z knihy Evolution of Man. Kniha 1. Opice, kosti a gény autora Markov Alexander VladimirovichNové gény? Génová aktivita sa počas vývoja môže meniť nielen zmenami v miestach viazania TF, prácami TF alebo regulačných RNA, ale tiež v dôsledku duplikácie génov. Keď sú iné veci rovnaké, dva identické gény budú produkovať viac produktu (t. J. Informácie
autor Harmar Hillery Z knihy Chov psov autor Harmar Hillery autor Harmar HilleryRecesívne gény Rekresívny gén (tj vlastnosť, ktorú určí) nemôže byť zobrazená jednou alebo mnohými generáciami, kým sa nedosiahnu dva identické recesívne gény od každého z rodičov (náhle prejavy tejto vlastnosti u potomkov by sa nemali zamieňať s
Z knihy Psy a ich chov [Chov psov] autor Harmar HilleryDominantné gény Prítomnosť dominantného génu sa vždy jasne a zvonka prejavuje zodpovedajúcim znakom. Preto dominantné gény, ktoré majú nežiaduci znak, predstavujú pre chovateľa oveľa nižšie riziko ako recesívne, pretože ich prítomnosť
Z knihy Psy a ich chov [Chov psov] autor Harmar HillerySmrteľné gény Ide o gény, ktoré spôsobujú smrť organizmu skôr, ako dosiahne pubertu. Smrteľné gény sú recesívne, tu sú niektoré príklady prejavu ich vplyvu: "rozštiepené pery a rozštiepené podnebie" je defektom v rozvoji hornej čeľuste, hemofília je nedostatok krvi
Z knihy Neandertálci [História frustrovanej ľudskosti] autora Vishnyatsky Leonid Borisovich autora Kandel Eric Richard Z knihy "Hľadanie pamäte" [Vznik novej vedy o ľudskej psychike] autora Kandel Eric Richard autora Hengstschleger MarkusGény športovec Ženy ho milujú za jeho vzhľad a muži majú radi svoju schopnosť získať lopty. Ženy obdivujú jeho účes a lákavý úsmev, muži ho rešpektujú za to, že ho vyhodili z boku. Je to asi dvaja ľudia. Nakoniec sme sa dozvedeli o genetike
Z knihy "Síla génov" (krásna ako Monroe, inteligentná ako Einstein) autora Hengstschleger MarkusMozgartove gény O Mozartovom detstve nie sú veľmi presné informácie, oveľa viac - predpoklady. Ako sa to malý génius stalo? Sestra Nannerl, o päť rokov staršia, sa chcela dozvedieť viac o svojom mladšom bratovi. Čo sa naozaj páčil v ranom detstve?
Z knihy "Síla génov" (krásna ako Monroe, inteligentná ako Einstein) autora Hengstschleger MarkusGENICKÉ GENE A teraz je úplne iná otázka: prečo je to tak zaujímavé? Čo môžete zistiť preskúmaním Mozartovej lebky? Nuž, povedzme, že bol zvážený a meraný. Podľa výsledkov analýzy kostí môžete zistiť, čo majiteľ tejto lebky jedol. A to už bolo urobené. Táto lebka
Z knihy Inner Fish [História ľudského tela od staroveku až po súčasnosť] autor Shubin NilGény Objavenie Arendtu nás prináša k ďalšej otázke. Je to jedna vec, že oči rôznych zvierat majú spoločné časti, ale ako sa stalo, že takéto odlišné oči ako červy, muchy a myši sú vzájomne spojené? Na odpoveď na túto otázku sa obrátime
Všeobecné údaje
Výsledky interakcie génov dvoch rodičovských genómov v zygote a mnohobunkovom organizme, ktoré sa z neho vyvíjali, sa prejavujú vo vlastnostiach, ktoré kontrolujú, ktoré sa do istej miery prenášajú z generácie na generáciu. Takáto dedičnosť závisí od mnohých dôvodov.
Po dlhú dobu v klasickej genetike sa predpokladalo, že príspevok oboch rodičov k genómu potomstva bol približne rovnaký pre materskú i otcovskú rodinu rodokmeňov rodokmeňov. Na tomto základe formulovali jedno z prvých dedičských pravidiel - rovnocennosť a prepojenie funkcií dvoch alel rôzneho pôvodu tých istých génov (alebo rovnocennosť vzájomných krížov).Identifikované dva varianty a niekoľko typov dedičnosti založené na počte génov, ich pôvod (otcovská materská alebo), lokalizácia v nealelickém loci alebo alely autozomů a pohlavných chromozómov, zobrazuje znak (dominantný alebo recesívny) a vlastnosti (mechanizmy) interakcia medzi génmi.
Avšak v druhej polovici dvadsiateho storočia že v mnohých prípadoch je príspevok jedného rodiča výrazne odlišný od príspevku druhého rodiča. Ukázalo sa, že funkcie rodičovských génov sa môžu meniť v priebehu celej ontogenézy až po diferencované odstavenie materských alebo otcovských alel. Základom tohto javu je epigénómový proces alebo označenie chromozómových lokusov jedného z rodičov, čo vedie k odstaveniu expresie alel nachádzajúcich sa v nich. Tento jav sa nazýva potlač.Termín bol prvýkrát použitý v druhej dekáde 20. storočia. Rakúsky zoolog Konrad Lorenz. Sledujúc správanie kačic, ktoré sa práve vyliahli z vajec, všimol si, že hľadajú vlastnú matku kačicu. Ak ich majiteľ alebo hosteska kačice alebo pes alebo mačka, ktorá prechádza okolo, príde do ich pohľadu, káčatko nasleduje celý svoj budúci život ako svoju matku.
Hovoríme teda o formácii pred narodením množstva funkcií tela, ktoré sú vytlačené v genóme ("genómová pamäť").
Neskôr termín "imprinting" začal používať neuropatológovia. Imprinting Sú presvedčení, že je v dôsledku vzniku mnohých vrodených javov v novorodenca, ako je hľadanie matky prsia bradavky alebo bázických funkcií (prehĺtaní, dýchanie, žuvanie, krvný obeh, trávenie a sanie), ktoré sú vytvorené počas fetálneho života.
Tento termín začal používať genetika na vysvetlenie nezvyčajného správania sexuálnych chromozómov (odstránenie chromozómov otcovského pôvodu) u hmyzu rodu Sciara Coprophila (G. Crouse, 1960). Bolo ukázané, že otcovské X chromozómy sú nejako označené (potlačené) pred zlúčením gamét v súlade s ich rodičovským pôvodom.
V súčasnosti je presvedčivo dokázaný všeobecný biologický význam imprintovania ako epigenomického procesu spojeného s regulačnými (funkčnými) zmenami v aktivite genómu a nesúvisiacich s jeho štrukturálnym poškodením (konv.
U ľudí, ako výsledok imprintovania, sa exprimuje materský gén (alela), zatiaľ čo otcovská alela je blokovaná alebo naopak. Z toho vyplýva, že jedinec má iba monoalický výraz alebo neefektívny príspevok k genómu jedného z jeho rodičov, čo znamená, že existuje odchýlka od Mendelovských zákonov o dedičstve založených na paralelnom modeli (pozri kapitolu 2).
Pred zvážením takýchto odchýlok sa budeme zaoberať vzormi dedičnosti génov a vlastností, ktoré najprv opísal G. Mendel na základe výsledkov jeho experimentov na prechode semien záhradného hrášku. V budúcnosti sa tieto vzory nazývajú zákony dedičnosti a rozšírenia na ľudí.
Zákony dedičnosti
V modernom vydaní sú zákony dedičnosti formulované nasledovne.
Prvý zákon- zákon o nadvláde (alebo jednotnosti) znaku v potomkoch prvej generácie. V prvej generácii prejavuje potomok pôsobenie dominantného génu (dominantný
symptóm), ale účinok recesívneho génu sa neprejavuje (recesívny znak). V ďalších generáciách potomok prejavuje pôsobenie dominantného aj recesívneho génu.
Druhé právo- zákon o štiepení génov u potomstva alebo zákon o "čistote gamet" V potomstve sa v gétách vyskytuje alternatívne rozdelenie (rozdelenie) génov: jedna polovica gamét nesie dominantné gény (A) a druhá polovica nesie recesívne gény (a). Obidva typy génov sa vyskytujú v somatických bunkách, nie v zmiešaní a ne nahradzovaní.
Tretí zákon- zákon nezávislé dedičstvo nealetických génov alebo náhodných kombinácií dedičných sklonov u potomkov. Združuje sa dedičnosť dvoch génov (dva páry znakov) dvoch-hybridný,viac ako dva gény (dva páry znakov) - poligibridnym.Vzorec pre takúto dedičnosť zodpovedá binomickej sérii: (3 + 1) n, kde η je počet génov (párov znakov). Vyvodiť vzorec použitý gril R. Pennet. Svojou pomocou sa vypočítajú genotypy organizmov, ich počet a závislosť od typov gamét obsahujúcich dominantné a recesívne gény (tabuľka 3).
Tabuľka 3.Lattice R. Penneta
Poznámka.A, B, A "a B" - dominantné gény; a, b, a "a b" sú recesívne gény; A "A, B" B, A "B a B" A - dominantné homozygotných organizmov; a "a, a" b, b "a, b" b - recesívne homozygotné organizmy; A "a, a" b, a "A, a" B, B "a, B" b, b "a b" B - dominantné heterozygotné organizmy.
Okrem troch zákonov dedičnosti určuje dedičnosť génov a postáv hlavné ustanovenia chromozomálnej teórie dedičnosti.
Chromozómová teória dedičnosti
V druhej dekáde XX storočia. Thomas Hunt Morgan (1866-1945) a jeho študenti (C. Bridges, G. Meller a A. Stertevant) formulovali hlavné ustanovenia chromozomálnej teórie dedičnosti(všetko
Moderná chromozómová teória dedičnosti zahŕňa nasledujúce ustanovenia.
Gény sa nachádzajú v chromozómoch; počet génov v chromozóme je úmerný dĺžke chromozómu. Neskôr sa ukázalo, že rôzne gény sa líšia, takže tento pomer nie je vždy rešpektovaný. Napríklad na chromozómoch 5 a 9 sú gény identifikované viac ako na najdlhších chromozómoch 1 a 2.
Gény sa nachádzajú pozdĺž dĺžky chromozómu lineárnym spôsobom. Následne boli identifikované mobilné genetické prvky alebo "skokové gény" (transpozóny), ktoré sa pohybujú pozdĺž genómu z jedného chromozómu na druhý a narúšajú lineárne usporiadanie.
Alelové gény obsadzujú identické lokusy homológnych chromozómov. Z dvoch homológnych chromozómov pôvodu je jeden otec, druhý je matka. Ich lokusy sú navzájom identické. Obsahujú alely toho istého génu alebo alelických génov. Každá z dvoch alel toho istého génu je otcovská a materská kópia; spravidla je to spravidla dielálny model organizačnej štruktúry génu (pozri obrázok 12). V prípade dedičnej patológie môže jednotlivec mať len jeden génový lokus, a preto bude existovať jedna alela (buď otcovská alebo materská); tento monoalelačný stav tejto alely je zaznamenaný napríklad v systéme Shereshevsky-Turnerovho syndrómu (karyotyp: 45, XO). Ďalším patologickým variantom je prítomnosť jedinca v rovnakom čase tri alely a viac; toto je stav polyallelizmu pre túto alelu - napríklad tri identické alely toho istého génu troch chromozómov 21 v Downovom syndróme (karyotyp: 47, XX, + 21); tri alely troch chromozómov 13 v Patauovom syndróme (karyotyp: 47, XY + 13) alebo prítomnosť karyotypu 4 až 11 X chromozómov (4-11 alel rovnakého génu - 49, XXXX).
Gény toho istého chromozómu tvoria väzbovú skupinu, ktorá zabezpečuje spoločné dedičstvo vlastností, ktoré kontrolujú. Treba poznamenať, že prepojenie génov s chromozómom je neustále narušované počas prechodu - procesom homológnej rekombinácie alebo výmenou identických oblastí (gény a ich fragmenty
policajti) medzi homológnymi chromozómami v prvej divízii meiózy. Frekvencia prekračovania je priamo úmerná vzdialenosti medzi génmi. Vzájomná výmena alel otcovského a materského pôvodu sa vyskytuje medzi všetkými pármi homológnych chromozómov, s výnimkou chromozómov X a Y chromozómov. Otvorenie prechodu tiež patrí do školy T.Kh. Morgan. Teraz zvážte možnosti a typy dedičnosti génov a vlastností.
MOŽNOSTI A TYPY VÝSKUMU GENE A SIGNS
V súčasnosti existujú tri varianty dedičnosti génov a znakov: monogénne a polygénne s tradičným (klasickým) dedičstvom a non-klasickým alebo netradičným dedičstvom.
Monogénna dedičnosť je založená na prvom a druhom zákone dedičnosti. Zahŕňa dedičnosť jedného génu (jeden pár znakov) a vzťahuje sa na alelické gény.
Polygénna dedičnosť je založená na treťom práve dedičnosti. Zahŕňa dedičnosť dvoch génov (párov znakov) a ďalšie a vzťahuje sa na nealetické gény.
Netradičná dedičnosť je dedičnosťou génov a znakov, ktoré idú nad rámec monogénnych a polygénnych variantov.
Monogénna dedičnosť
Monogénne dedičstvo sa často nazýva jednoduché mendelovské dedičstvo.
Na základe myšlienok o diallelickom modeli štruktúry génu sa dedičstvo interakčných mechanizmov medzi otcovským a materským genómom zvažuje zvlášť pre každý alelický a nealový pár.
Druhy monogénnej dedičnosti
V rámci monogénneho dedičstva existujú:
Autozomálny dominantný typ (dominantný gén sa nachádza na jednom z dvoch autozómov);
autozomálne recesívny typ (recesívny gén sa nachádza na jednom z dvoch autozómov);
X-viazaný dominantný typ (dominantný gén sa nachádza na X chromozóme);
X-viazaný recesívny typ (recesívny gén sa nachádza na chromozóme X);
Y-viazaný typ alebo holandrická dedičnosť (gén sa nachádza na chromozóme Y).
Podľa katalógu V. McKusicka "Dedičnosť mendelovských znakov u ľudí" (internetová verzia: online - http: www.ncbi.nlm.Nih.gov/Omim) bolo identifikovaných viac ako 12,5 tisíc takýchto fenotypov. Medzi nimi približne 12 tisíc fenotypov zdedených (alebo pravdepodobne zdedených) autozomálne dominantne alebo autozomálne recesívne (vrátane 9 tisíc fenotypov so zavedeným typom dedičnosti).
V posledných rokoch bolo získaných množstvo údajov o dedičstve spojených s chromozómami X a Y, na ktorých sú lokalizované.
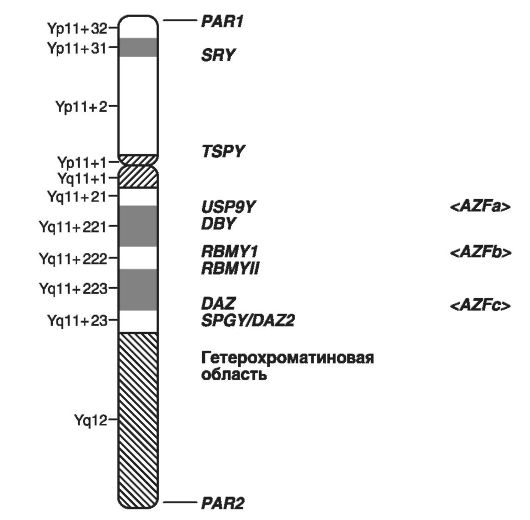
300 a 92 génov.
Na obr. Obrázok 20 znázorňuje mapu chromozómu X a množstvo dominantných a recesívnych ochorení spojených s jeho lokusmi.
Nasledujúce zriedka sa vyskytujúce ochorenia by sa mali uviesť ako príklady dominantných fenotypov viazaných na X: rachety alebo hypofosfatémia odolné voči vitamínu D (Hr22.2); syndróm inkontinencie pigmentov, typ I (Xp11.1) a typ II (Xq28).
Príklady najbežnejších chorôb dedičného X-viazaná recesívne: hemolytická anémia (Xq21.2 alebo Xq28), hemofília A (Xq28.2) a B (Xq27.2), Duchennova svalová dystrofia, Becker (Xp21.2), syndrómy Lesch -Nayana (Xq26.2),
bodová kondysplázia Konradi-Hünermanna (Hr22.2), retinitis pigmentosa (Xp21.2-21.3, Xp22).
Na obr. 21 znázorňuje schematické znázornenie lokusov chromozómov Y, v ktorých sú umiestnené gény tvoriace mužský pohlavie a gény spôsobujúce mužskú neplodnosť.
Ako príklady chorôb spojených s chromozómu Y by mala viesť k diferenciácii porušeniu podlahovej formy mužskej neplodnosti je azoospermia (YP - faktor 2; Yq11 - faktor 1)., Gonadoblastomu atď. Celková schéma izolovaných génov zodpovedných za siedmich z týchto chorôb ,
Okrem toho, monogénne dedičstvo môže byť nielen spojená s podlahou (X alebo Y chromozóm), ale tiež obmedzená na podlahu.
Obr. 20.Mapa ľudského X chromozómu (podľa multimediálnej učebnice "Biológia" Copyright (c), 2007)
Napríklad, gén plešatosti sa prejavuje u mužov (dominantný účinok) a takmer sa neprejavuje u žien (recesívny účinok).
Kritériá pre monogénne dedičstvo
Autozomálny dominantný typ:
Toto ochorenie sa pravidelne prenáša z generácie na generáciu bez medzery, t.j. sledované v rodokmeni vertikálne, okrem prípadov de novo mutácií (pozri kapitolu 5);
Obr. 21.Schematické znázornenie chromozómu Y (podľa GenBank, 2003)
Riziko mať choré dieťa, ak je jeden z rodičov chorý, je 50%;
Zdraví jedinci majú zdravé potomstvo;
Nemocný pacient má jedného z rodičov okrem prípadov novej mutácie;
Obidve pohlavia sú postihnuté rovnakou frekvenciou. Autozomálny recesívny typ:
Rodičia probandového pacienta (osoba, ktorá konzultovala s genetickým lekárom) sú zdravé, ale podobná choroba sa vyskytuje u príbuzných probandov, bratrancov a druhých bratrancov (jeho bratov a sestier), t. J. sledované horizontálne v rodokmeni (v jednej generácii);
Chorý rodič porodí zdravé deti;
Riziko mať choré dieťa je 25% (pomer chorých a zdravých ľudí je 1: 4);
V prípade dedičných manželstiev medzi rodičmi probandujúceho pacienta sa pozoruje nárast počtu chorých príbuzných v rodokmeňoch.
X-viazaný dominantný typ:
Probandový pacient nevyhnutne má jedného rodiča;
Chorý otec má všetky svoje dcéry choré a jeho synovia sú zdraví;
Nemocná matka má rovnako pravdepodobne chorú dcéru a chorého syna;
So zdravými rodičmi budú všetky deti zdravé;
Existujú 2 krát viac chorých žien ako chorých mužov. X-viazaný recesívny typ:
Choroba sa pozoruje u mužských príbuzných probandu pacienta na materskej línii;
Synovia nezdieľajú chorobu otca;
V chorom otcovi sú všetky dcéry zdravé a sú heterozygotné nosiče génu otcovskej choroby;
Ak je žena heterozygotným nosičom génu ochorenia, polovica jej synov bude chorá a všetky dcéry sú zdravé; a polovica dcérov sa stane tiež heterozygotnými nosičmi génu choroby.
Kritériá holandského dedičstva ešte neboli vyvinuté. V posledných rokoch bolo u ľudí zistené množstvo zložitých zdedených monogénnych a polygénnych ochorení:
Choroby tráviaceho ústrojenstva - rakovina prsníka (dve genotypy: gény VPC1 a VRC2);
Trigénne choroby - Bardet-Beadleov syndróm (3 genocídy: gény BBS1, BBS2 a BBS30);
Kvadrogénne pentagénne ochorenia: Alzheimerova choroba (spôsobená štyrmi genocípami: PS1, PS2, PS3 a PS4, rovnako ako genóm priónového proteínu, pozri kapitolu 29);
Polygénne alebo multifaktoriálne ochorenia (MDF) spôsobené "génovými sieťami" - arteriálna hypertenzia (170 génov vrátane 17 hlavných génov), bronchiálna astma (asi 20 génov), osteoporóza (13 génov), endometrióza (11 génov) atď.
Polygénna dedičnosť
Polygénna dedičnosť sa často nazýva multigénom alebo multifaktoriálnym, čo znamená dedičnosť v tom istom čase nie jedného, ale niekoľkých špecifických génov, ktoré prejavujú ich
pôsobenie v špecifických podmienkach prostredia za prítomnosti provokujúcich vonkajších faktorov, spravidla zvyšujúcich individuálny účinok génov, ktorých účinok je kumulatívny (aditívny účinok).
Kritériá polygénneho dedičstva (5 celkovo) boli systematizované v roku 1969 K. Carterom a 20 rokov neskôr boli doplnené (ďalšie dva) F. Vogelom a A. Motulskim (1989).
Polygenetické kritériá dedičnosti
Riziko vzniku multifaktorového znaku (ochorenia) je určené nasledujúcimi faktormi.
Dedičstvo znaku alebo choroby. Čím vyššia je heritabilita znaku alebo choroby (čím viac zodpovedajú dedičné gény), tým väčšie je riziko jeho vývoja u zdravých príbuzných. dedičnosťznamienko (choroba) je stupeň vplyvu na vznik daného znaku (ochorenia) dedičných faktorov v porovnaní s faktormi životného prostredia. Dedičnosť sa vyjadruje v absolútnych číslach (od nuly do jedného) alebo v percentách s použitím koeficientu h 2 alebo K n, ktorý sa vypočíta podľa vzorca: K n = G / E x 100%, kde K n je dedičský koeficient, G je dedičné faktory, E - environmentálne faktory. Tabuľka 4 zobrazuje hodnoty počtu faktorov dedičnosti multifaktoriálnych znakov a chorôb.
Tabuľka 4.Koeficienty dedičných multifaktoriálnych znakov a chorôb
Závažnosť symptómu alebo závažnosť ochorenia v probande. Čím silnejšie je znamenie alebo ťažšie úniku
ochorenie u chorého príbuzného, tým vyššie je riziko jeho vývoja u zdravých príbuzných.
Zvyčajnosť génov v probande a jeho príbuzných (alebo blízky stupeň príbuzenstva s chorým príbuzným). Častejšie gény u pacienta a jeho príbuzných, tým vyššie je riziko rozvoja znaku alebo choroby v druhom. Napríklad frekvencia populácie psoriázy je 0,75%. Medzi príbuznými prvého stupňa príbuzenstva je frekvencia ich vývoja 5,6%, medzi príbuznými druhého stupňa príbuzenstva - 3,0-3,5%, medzi príbuznými tretieho stupňa príbuzenstva - 1,75%, medzi príbuznými štvrtého stupňa príbuzenstva - 0,75% ,
Zriedka postihnutý sex. Multifaktoriálny znak alebo choroba sa prejavuje častejšie u jedincov zriedkavo postihnutého pohlavia (kritérium nazývané carterov efekt).Napríklad vrodená pylorická stenóza sa u chlapcov vyskytuje 2-5 krát častejšie ako u dievčat, t.j. v tomto prípade je ženský pohlavie zriedka postihnutým pohlavím. Avšak frekvencia tohto ochorenia u budúcich detí dievčat postihnutých pylorickou stenózou dosiahne 10-20%, zatiaľ čo v budúcnosti deti chlapcov postihnutých pylorickou stenózou - len 2-6%. Ďalším príkladom je žalúdočný vred a dvanástnikový vred, ktorý sa spravidla prejavuje u mužov a oveľa menej často u žien. Avšak jeho frekvencia u detí chorého je vyššia ako u detí chorého človeka.
Počet chorých príbuzných. Čím viac príbuzných v rodokmeni majú viacfaktorový znak alebo chorobu, tým väčšie je riziko jeho vývoja medzi potomkami (tabuľka 5).
Tabuľka 5.Riziko vývoja multifaktorového znaku alebo choroby v probande v závislosti od počtu jeho chorých príbuzných
Celkové riziko pre deti, ak sú rodičia zdravé, je 5 - 10%; ak je jeden z rodičov chorý - 10-20%; ak sú obaja rodičia chorí - až 40%;
Ďalšie kritériá
Dvojité kritérium. Ak je zhoda (podobnosť) monozygóznych dvojčiat na niektorom znamení alebo ochorení je 4 krát vyššia ako zhoda s dizygotnými dvojčatami, potom sa tento príznak alebo choroba zdedí podľa polygénneho variantu.
Kritérium segregácie postihnutých súrodencov v rodinách s jedným chorým alebo dvoma zdravými rodičmi. Ak je podiel chorých súrodencov v rodinách s jedným chorým rodičom 2,5-krát vyšší (a viac) ako podiel chorých súrodencov v rodinách s dvoma zdravými rodičmi, je pravdepodobný najmä polygénny variant dedičnosti. Ak je tento pomer menší ako 2,5, potom nie je vylúčená aj polygénna dedičnosť.
Avšak klasické varianty dedičnosti génov a vlastností sa vyskytujú len v menšej časti všetkých prípadov takej dedičnosti, zatiaľ čo takmer dve tretiny z nich sú netradičné varianty.
Netradičné dedičstvo
Netradičné dedičstvo je zaznamenané v mnohých dedičných chorobách človeka. V posledných rokoch boli im pridelené predtým slabo študované alebo všeobecne neznáme choroby šiestich tried: akumulačné ochorenia (44 nosopisov, pozri kapitolu 21); peroxisomálne ochorenia (17 nosológií, pozri kapitolu 26); mitochondriálne ochorenia (rozlišovalo sa viac ako 200 nosológií, predpokladá sa ďalších 1150, pozri kapitolu 26); impresionistické choroby (zvýraznené sú 24 nosopisov, pozri kapitolu 28); ochorenia rozšírenia počtu nukleotidových opakovaní (27 nosopisov, pozri kapitolu 27); priónové ochorenia (pozri časť 29).
Vo všeobecnosti sú netradičnými chorobami v dedičstve 321 nosologických jednotiek (okrem 1 150 mitochondriálnych ochorení). Z dedičstva chorôb prvých troch tried je to asi materskej dedičnostipretože lysozómy, mitochondrie a peroxizómy, ktoré sú cytoplazmatickými štruktúrami somatických buniek, sa dedia výlučne cez linku matka a dcéra a nikdy sa neprejdú pozdĺž matky-syna, otca-syna alebo otcovo-dcérnej línie, čo je spôsobené biologická matriarchia(pozri kapitoly 5 a 12).
Avšak pri mnohých mitochondriálnych ochoreniach, okrem čisto materskej dedičnosti, možno pozorovať monogénnu dedičnosť zmiešanú s matkou. Je spojená so zhoršenou interakciou medzi mitochondriálnymi (mtDNA) a jadrovými (DNA) genómami. Najmä gény kódujúce mitochondriálne proteíny sa nachádzajú buď v mitochondriách (gény dvoch typov rRNA, 22 typov tRNA, 13 typov polypeptidov), alebo v bunkovom jadre (existuje okolo 1150). Jadrové gény sú transkribované v jadre, preložené v cytoplazme a výsledky ich expresie sú "importované" mitochondriami.
Obvykle sú všetky kópie mtDNA navzájom identické (homoplázia). V prípadoch, keď sa vyskytujú mutácie v mtDNA (ich frekvencia je 10 krát častejšia ako v jadrovej DNA), dva typy mtDNA sú možné v jednej bunke (heteroplázia) a potom sa nazýva proces šírenia mutantnej a normálnej mtDNA replikačná segregácia(pozri kapitolu 2).
V prípade, že potlač ochorenie alebo epigenetických vypnutie expresie loci chromozómov rodičov (pozri vyššie). Fenotyp gén akcie sa môžu meniť v dôsledku troch faktorov: delécie génu (genetické kontaktných platní) uniparentální izodisomiya (chromozómové potlač) a narušenie génovej expresie v srdcovej imprinting ,
Genetická imprintingzaložené na mechanizme špecifickej metylácie cytozínových báz molekuly DNA, ktorá vypne génovú transkripciu (pozri kapitolu 28).
V súčasnej dobe sa predpokladá existenciu asi 200 génov, a náchylný k takej potlačená monoallelic expresiu tkanivovo špecifických, ako aj tri génového klastra nájdených lokalizované v kritických oblastiach chromozómov (7q32, 11p15 a 15q11.2-Q13), a majú priamy vplyv na rozvoj dedičných chorôb takýto mechanizmus vrátane nádorov.
Genómová imprintingzaložené na homogénnej izodizóme na špecifickom chromozóme materského alebo otcovského pôvodu. Pozoruje sa v mnohých dedičných a vrodených ochoreniach, ako sú: Angelmanove syndrómy (spôsobené homogénnou isodizómiou na otcovskom chromozóme 15) a Prader-Willi (izodizómia 15 materských chromozómov); prechodné neonatálne diabetes mellitus (otcovský chromozóm 6); Syndróm Silver Russell (materský chromozóm 7); syndrómu
Beckwith-Wiedemann (otcovský chromozóm 11); oneskorený fyzický a motorický vývoj, hypotenzia a predčasný pohlavný vývoj (materský chromozóm 14); intrauterinná retardácia rastu (IUGR) plodu (materský chromozóm 7), rovnako ako IUGR s obmedzeným placentovým mozaicizmom (materský chromozóm 16).
Účinok genomickej imprintáže sa vyskytuje aj počas bublinovej šmykovej dráhy. Existujú 4 typy porušení:
Androgenéza alebo dvojitá sada chromozómov (2n): dve spermie s X chromozómami a jedno vajíčko bez jadra;
Genogenéza alebo dvojitá sada chromozómov (2n): bunka vajíčka s dvojitou sadu chromozómov, spermie nie sú zapojené do oplodnenia;
Android alebo trojnásobná sada chromozómov (3n): 2 otcovské a 1 materská;
Ginoidná alebo trojnásobná sada chromozómov (3n): 2 matky a 1 otcovská.
Impressum v dôsledku chýb génovej expresie v imprinte
sa vyskytuje aj u Angelmanových a Prader-Williových syndrómov. Je to spôsobené tým, že obaja rodičia prenášajú na potomka pacienta gény, ktoré nesú svoje špecifické vlastnosti, t.j. gény otca a matky sú aktivované alebo potlačené potomkami rôznymi spôsobmi (tzv gény vytlačené).Navyše, v rade matky i otca, sú to parentálne poruchy génovej expresie, ktoré sú zdedené v imprinte.
V súčasnosti bolo na 1, 5, 7, 11, 13, 15, 19 a 20 izolovaných najmenej 60 odtlačených génov (alebo ich transkriptov) izolovaných a X chromozóm.
Treba poznamenať, že v systéme Angelman a Prader-Willi boli nájdené aj úzko lokalizované, ale opačne označené gény. Boli povolaní kandidátskych chorobných génov(v rodinách s opakovanými prípadmi týchto syndrómov). A genykandidaty Angelmanův syndróm sú exprimované výlučne na materskej chromozóme, ale potlačila otcovská chromozóm, zatiaľ čo genykandidaty Prader-Williho syndróm sú exprimované na otcovskej chromozóme, ale potlačila matiek chromozóm (pozri kap. 28).
Imprintovanie je výsledkom kvalitatívnych, nie kvantitatívnych zmien dedičného materiálu.
Na konci XX storočia. tiež identifikovala predtým neznámu triedu chorôb s netradičným dedičstvom - expanzné ochorenia
týždňové opakovaniaspojené so zvýšením počtu kódovacích a nekódujúcich nukleotidových sekvencií v dôsledku dynamických mutácií. Dynamická mutácia sa "pohybuje" zo stavu fenotypicky neprejavenej premutáže na stav fenotypicky manifestovanej plnej mutácie. Takéto mutácie sú základom mnohých závažných dedičných neurodegeneratívnych ochorení (27 nosopisov, pozri kapitolu 27). Napríklad opakovanie CGT je typické pre Martin-Bell syndróm (Xq23), opakovanie GHZ pre druhú variantu tohto syndrómu (Xq28); CTG opakovanie - pre spin-bulbar svalovú atrofiu (Xq11-12); CAG - pre myotonickú dystrofiu (19q13.3); CTG pre Huntingtonovu chorea (4p16.3); CTG - pre spinocerebrálnu ataxiu typu I (6p21.3); CTG - pre Machado-Josephovu chorobu (14q32.1); TTC - pre Friedreichovu ataxiu (9p13). Vo všetkých prípadoch dochádza k postupnej akumulácii (rozširovaniu) kritického počtu nukleotidových opakovaní.
Súbežne s expanziou v každej nasledujúcej generácii sa zvyšuje závažnosť ochorenia - predvídavosť.
INDIKÁTORY GENOVÝCH INTERAKCIÍ AKO DOPLNKOVÉ KONCEPCIE GENOMIKY A PROTÓNY
Podľa zákonov genetiky sa otcovské a materské genómy, spojené v genotype jednej bunky a celého organizmu, vzájomne vzájomne ovplyvňujú počas ontogenézy, vytvárajúc individuálnu vonkajšiu a vnútornú charakteristiku, fenotyp organizmu.
Existujú tri skupiny mechanizmov interakcie medzi rodičovskými génmi a podľa toho tri skupiny ukazovateľov súvisiacich s ďalšími koncepciami genomiky a proteomiky. Ide o interakcie medzi:
Alelické gény; charakterizuje ich: dominancia, recesia, dominancia, neúplné a podmienené dominantné postavenie, naddomínstvo;
Nealetické gény; sú charakterizované: epistázou, komplementaritou, polymérmi;
Oddelený genóm a genotyp (ako systém génov); sú charakterizované: expresivitou, penetráciou, účinkom polohy,
genocída, pleiotropický efekt, alelické série a fenokopy.
Zvážte tieto mechanizmy ako dodatočné koncepty genomiky a proteomiky.
Mechanizmy interakcie medzi alelickými génmi
Dominovanie a zapadanie
Ak funkčný stav jednej alely (napríklad materskej alely) nezávisí od stavu druhej alely (otcovskej alely), potom potomok bude vykazovať znak ovládaný materskou alelou. Takýto gén a jeho znak sa nazývajú dominatnymi.
Prvý popis účinku dominancie sa týka roku 1905, keď bola rodina pacienta s brachydactyly označená krátkou päsťou. Ďalšími príkladmi sú: biele vlnenie, nočná slepota"Habsburg ret polydaktýlia (polydactyl) syndaktylia (fúzie mäkkých tkanív alebo kostí prstov) arachnodactyly (" pavúčie prsty ") chondrodystrophy a početné formy autozomálne dominantné ochorenie, ako je napríklad Huntingtonova chorea (4r16.3).
Ak funkčný stav jednej alely (napríklad materskej alely) závisí od stavu druhej alely (otcovskej alely), potom potomka prejaví znak, ktorý je riadený oboma materskými a otcovskými alelami súčasne. Súčasne sa nazýva aj samotný gén, ako aj vlastnosť, ktorú kontroluje recesívny.
Príklady dedičnosti recesívnych znakov a fenotypov: albinizmus, mäkké, rovné vlasy, strnulý nos, jasné oči, rhesus-negatívny krvný typ I, neschopnosť cítiť chuť fenyltiokarbamidu; početných foriem autozomálnych recesívnych ochorení, ako je genokopia fenokonónírie (12q24.2, 4p15.1).
V niektorých prípadoch dominancia a recesivita génov koreluje slabo s dominanciou a recesívnosťou postáv. Napríklad mongoloidný epikant je riadený dominantným génom a Bushmenom a Hottentotom, recesívnym génom.
Ďalším príkladom je uvedený gén a znak plešatosti, ktorý sa prejavuje u mužov ako dominantný a u žien ako recesívny. Ako sa ukázalo, takýto mechanizmus je spôsobený pôsobením hormónov, t.j. existuje rodovo závislé (sex riadené) dedičstvo.
Neúplné dominantné postavenie
O neúplnej dominancii,alebo prechodné pôsobenie génov (prejav príznakov), vyslovujú sa pri oslabení pôsobenia dominantného génu v prítomnosti recesívneho génu, t.j. v heterozygotoch. Nemožno však vyvodiť jasnú hranicu medzi prechodnými opatreniami a dominantným postavením na jednej strane, ako aj prechodnými opatreniami a recesiou na strane druhej. Napríklad, pigmentácia ľudskej kože sa líši od bielej v albino po čiernu v čiernom. Od sobášov medzi bielymi a čiernymi sa narodili mulatovia, ktorí majú strednú farbu kože.
Ďalším príkladom neúplnej dominancie sú rozdiely v 6 druhoch speváckych hlasov, ktoré sú riadené jedným alelickým párom. Najmä baryton a mezzo-soprán sa pozorujú len u heterozygotov, zatiaľ čo tenor a bas, alto a soprán sú charakteristické pre homozygoty.
V ďalších štúdiách sa však zistilo, že pigmentácia kože a druh hlasu spevu určujú nielen tento mechanizmus interakcie, ale aj faktory nezávislé od seba: vplyv pohlavných hormónov, vplyv polymérneho, pohlavne viazaného alebo pohlavne závislého dedičstva.
Podmienečné (nestabilné) dominantné postavenie
Keď nestabilné alebo podmienené nadvládyprejav znaku heterozygotov závisí od genotypu a vonkajších podmienok (modifikačný účinok genotypu na hlavný gén, penetrácia génu, umiestnenie génu v chromozóme, vplyv teploty).
Kodominirovanie
Ak sú alelické gény rovnako aktívne (majú rovnaké dominantné pôsobenie), potom toto kodominirovanie.Jeho klasickým príkladom je dedičnosť IV krvnej skupiny (podľa systému ABO), určená tromi alelami nachádzajúcimi sa v 9. chromozóme (množine alel). Medzi nimi sú dve dominantné alely (IA a B) a jedna recesívna alela (IO). Párna kombinácia týchto alel poskytuje 4 krvné skupiny:
- prvej skupiny- prítomnosť dvoch identických recesívnych alel - I010 (homozygot), určenie prítomnosti protilátok alfa a beta v krvnom sére;
- druhej skupiny- prítomnosť dvoch identických dominantných alel IAIA (homozygot) alebo dvoch rôznych alel IA10 (heterozygot), čo spôsobuje prítomnosť beta-protilátok v sére;
- tretia skupina- prítomnosť dvoch identických dominantných alel MB (homozygot) alebo dvoch rôznych alel M0 (heterozygot); alfa protilátky sú prítomné v krvnom sére;
- štvrtej skupiny- prítomnosť dvoch rôznych dominantných alel! A! B (heterozygot); v sére nie sú žiadne protilátky, obe alely interagujú s rovnakou silou a neutralizujú sa navzájom.
Ďalším príkladom dominancie je dedičnosť kosáčikovej anémie, ktorá je autozomálna recesívna porucha (11p15). V tomto prípade sa pozoruje homozygotnosť (2 patologické alely jedného génu riadiaceho syntézu defektného hemoglobínu). Takéto homozygóty majú charakteristické symptómy, ale sú imúnne voči malárii, pretože plazmodium malárie nie je reprodukované na defektnom hemoglobíne.
Avšak v heterozygotnom organizme sú súčasne prítomné normálne a defektné alely toho istého génu. Navyše obe alely dávajú rovnaký dominantný účinok, a preto sú v bunkách súčasne syntetizované dva typy hemoglobínu (normálne a abnormálne). Takéto heterozygotné nosiče patologického génu nemajú príznaky kosáčikovej anémie alebo sa prejavujú v miernom stave a len v podmienkach nedostatku kyslíka.
superdominancie
V niektorých prípadoch sú alely, ktoré sú v heterozygotnom stave, fenotypicky silnejšie než alely, ktoré sú v homozygotnom stave. (efekt predominancie).Takýto prejav sa podobá heterolisový efektv rastlinách (hybridná sila alebo sila). Takže v prípade manželstiev medzi zástupcami rôznych rás sú zdravotné ukazovatele ich potomkov nadradené zdravotným ukazovateľom samotných rodičov: deti majú vyššiu vitalitu, dlhšiu životnosť atď.
Mechanizmy interakcie medzi nealeálnymi génmi
epistáze
epistáze- potlačenie pôsobenia génu umiestneného v jednom neadekvátnom páre pôsobením génu z iného nealkoholového páru, napríklad supresiu génu A génu B, t.j. A\u003e B alebo A\u003e bb. Rozlišujte dominantnú a recesívnu epistázu.
Dominantná epistáza:dominantná alela jedného neeletického páru, ktorá je v homozygotnom (AA) alebo heterozygotnom (Aa) stave, potláča prejav nealeálnej dominantnej alely druhého alelického páru, ktorý je v stave AA alebo Aa. Dominantné gény sa nazývajú epistatických génovalebo supresorov (inhibítorov). Môžu byť dominantné aj recesívne. Potlačené gény sa nazývajú hypostatických génov.
Ak sa gény nachádzajúce sa v iných nealových pároch zvýšia dominantný účinok epistatických génov, potom sa nazývajú modifikujúce gény(Zosilňovače).
Tento typ interakcie je charakteristický pre nealetické gény, ktoré sa podieľajú na regulácii ontogenézy, napríklad gény imunitnej odozvy (génová sieť - 2190 gény, viď kapitola 15) alebo gény erytropoézy (génová sieť - 200 génov).
Existujú dve možnosti dominantnej epistázy:
Homozygoti s recesívnymi alelami (aa) sa líšia fenotypom od homozygotov s dominantnými alelami (AA);
Homozygóty pre dominantné alely (AA) sa nelíšia v fenotype z homozygotov pre recesívne alely (aa).
Recesívna epistázaže recesívna alela jedného génu potláča účinok nealylového dominantného génu (aa\u003e B) a komplementárnosť sa pozoruje medzi dominantnými génmi (pozri nižšie). Príklad recesívnej epistázy u ľudí je "fenomén Bombay" spojený s narodením detí s I (I 0 I 0) a IV (I A I B) krvných skupín od rodičov s I (I 0 I 0) a II (IAI) krv, zatiaľ čo teoreticky od týchto rodičov sa majú narodiť deti s I (I 0 I 0) alebo II (IAI 0) krvné skupiny. Tento jav možno vysvetliť buď prítomnosťou nie je platné v jednej nadradenej vzácnych heterozygotná forma III krvné skupiny (IBI 0) alebo v prítomnosti genotype dieťaťa s IV krvnej skupiny (I A I B) recesívnych modifikačné gény, ktoré sú homozygotná inhibujú expresiu antigénov erytrocyty umiestnené na povrchu, t.j. dávajú nepredvídateľný fenotypový účinok.
Okrem recesívnej epistázy je zvýraznené dvojitá recesívna epistáza;počas recesívnych génov má svoj vlastný fenotypový prejav av dvojitých homozygotiach sa recesívne alely navzájom potlačujú: aa\u003e bb, bb\u003e aa.
komplementarita
komplementarita- typ interakcie najmenej dvoch dominantných nealetických génov z niekoľkých párov s rôzne kombinácie dominantných a recesívnych alel, čo prispieva k vývoju iného znaku iného, ako sú rodičovské varianty.
Existujú tri typy komplementárnosti:
Dominantné alely (AB) sa líšia fenotypovým prejavom;
Dominantné alely (AB) sú podobné pri fenotypovom prejave;
V dominantných (A) a recesívnych (a) alelách niekoľkých nealetických párov existuje nezávislý fenotypový prejav.
Napríklad u osoby je normálne počutie spôsobené interakciou niekoľkých dvojíc nealkoholových génov, ale aspoň jedna dominantná alela musí byť prítomná v pároch. Ak sa človek ukáže ako recesívny homozygotný (aspoň jeden pár nealkoholových génov), potom bude hluchý.
Ďalšími príkladmi komplementarity sú fenotypy pacientov s rakovinou s retinoblastómom a nefroblastómom (pozri kapitolu 25).
Polymér a kvantitatívna charakterizácia
polymerism- to je podmienenosť znaku alebo fenotypu interakciou génov lokalizovaných v niekoľkých nealkoholických pároch a poskytujúcich rovnaký účinok. Tieto gény sa nazývajú polymérnych génovalebo polygénov. Stupeň prejavu znaku (fenotypu) závisí tak od počtu dominantných génov v nealkoholických pároch, ako aj od počtu nealitických párov.
Takéto označenia sa nazývajú kvantitatívne znaky.Výrazne sa líšia od kvalitatívnych vlastností.
Ak sú kvantitatívne znaky riadené polygénne zdedenými génmi a v mnohých štátoch sa vyskytujú ako prechodné formy, potom kvalitatívne znakysú riadené génmi zdedenými monogénne a vyskytujú sa iba v alternatívnych štátoch bez prechodných foriem.
Príklady kvantitatívnych znakov sú: vyššie uvedená zmena pigmentácie kože u ľudí a prítomnosť strednej farby kože v mulatách.
Ďalšími príkladmi kvantitatívnych znakov sú úroveň (stav) zdravia človeka, očakávaná dĺžka života, intelektuálne schopnosti, hmotnosť a dĺžka tela.
V posledných rokoch sa zdôraznil fenomén interakcie medzi početnými neleálnymi génmi: kumulatívnych polymérov.V tomto prípade hovoríme o aditívnej (sumárnej) akcii génov, z ktorých každá má svoj (často malý) účinok na znak. Je to kumulatívny polymér, ktorý tvorí vyššie uvedené génové siete, ktoré riadia významnú časť kvantitatívnych znakov organizmu.
Medzi génmi, ktoré ovplyvňujú kvantitatívny atribút, môže existovať jeden hlavný gén a niekoľko génov, ktoré sú v porovnaní s ním (polygény) slabšie. Účinok hlavného génu je niekedy výrazne vyšší ako účinok iných génov a znak, ktorý je riadený hlavným génom, sa zdedí ako Mendelovská (monogénna dedičnosť) a znaky riadené polygénmi sa dedia podľa polygénneho variantu. Príkladom je dedičstvo trpaslíkov spôsobené hlavným génom v prípade achondroplázie, zatiaľ čo u normálnej populácie je ľudský rast určený aditívnym účinkom polygénov.
MECHANIZMY INTERAKCIE MEDZI
SEPARÁTOVÝ GENE A GENOTYPE
Výraznosť a penetrácia
Tieto pojmy boli prvýkrát predstavené v roku 1926 N.V. TimofeevRessovskim a 0. Vogt, aby opísali rôzne prejavy znakov a génov, ktoré ich kontrolujú. expresivitaexistuje určitý stupeň manifestácie (variácie) rovnakého znaku u rôznych osôb s génom, ktorý kontroluje tento znak. Existuje nízka a vysoká expresivita. Zvážte napríklad rozdielnu závažnosť rinitídy (rýľový nos) u troch rôznych pacientov (A, B a C) s rovnakou diagnózou 0RVI. V prípade pacienta A sa rinitída prejavuje miernym stupňom ("čuchanie"), čo umožňuje, aby človek počas dňa manipuloval s jedným vreckom; pacient má rinitídu vyjadrenú v miernych (denne 2-3 vreckovky); Pacient C má vysokú závažnosť rinitídy (5-6 vreckovky). Keď hovoríme o expresivite nie je jediný znak, ale všeobecne choroby, lekári často posudzujú stav pacienta ako uspokojivý alebo mierny alebo ťažký,
tj v tomto prípade je pojem expresivity analogický s pojmom "závažnosť choroby".
penetrance- je to pravdepodobnosť prejavu rovnakého znaku u rôznych jedincov s génom, ktorý kontroluje tento znak. Penetrácia sa meria v percentách jedincov so špecifickým znakom z celkového počtu osôb, ktoré nesú gén, ktorý kontroluje tento rys. 0na je neúplná alebo úplná.
Príkladom ochorenia s neúplnou penetráciou je stále rovnaká nádcha s OPVI. Takže môžeme predpokladať, že pacient A nemá rinitídu (existujú však iné príznaky choroby), zatiaľ čo pacienti B a C majú rinitídu. Preto je v tomto prípade penetrácia rinitídy 66,6%.
Príklad ochorenia s plnou penetráciou - autozomálna dominantnosť chorea Huntington(4r16). Vykazuje sa najmä u osôb vo veku 31-55 rokov (77% prípadov), u ostatných pacientov v inom veku: tak v prvých rokoch života, ako aj v 65, 75 rokoch a viac. Je dôležité zdôrazniť, že ak sa gén tohto ochorenia prenesie na potomka od jedného z rodičov, choroba sa nevyhnutne prejaví, aká je úplná penetrácia. Je pravda, že pacient nie vždy žije s prejavom Huntingtonovej chorey, zomiera z inej príčiny.
Účinok polohy
Iný typ závislosti genu na genotype - pozitívny efekt.Otvoril ju A. Stertevant (1925). Podstata tohto účinku je v zmene expresie génu so zmenou jeho polohy (pozície) v chromozóme (v sérii nukleotidových sekvencií).
Podľa moderných konceptov nie je vplyv pozície spojený s porušením štruktúry génu: táto a jeho promótorová oblasť sú zachované ako jednotka transkripcie. V dôsledku toho je účinok pozície epigenomická udalosť určená troma podmienkami:
V promótore sa začína transkripcia;
Regulačné prvky (zosilňovače a tlmiče) obsahujúce väzbové miesta transkripčného faktora zvyšujú špecifickosť transkripčného komplexu na promótore;
Organizácia chromatínu v lokusovej oblasti prispieva k zvýšeniu citlivosti na pôsobenie nukleáz.
Úroveň expresie závisí od umiestnenia génu v genóme: buď v oblastiach kondenzovaného heterochromatínu, alebo v oblastiach
undekondenzovaný chromatín (euchromatín), ktorý je de-kondenzovaný v interfáze, obsahuje väčšinu génov a replikuje sa na začiatku S-fázy. Na druhej strane heterochromatín kondenzuje v celom bunkovom cykle, replikuje sa na konci S fázy a obsahuje väčšinou opakujúce sa sekvencie. Centromérne oblasti chromozómov pozostávajú zo štrukturálnych heterochromatínov alebo husto kondenzovaných chromatínov obsahujúcich opakujúce sa sekvencie.
Vzhľadom na tieto zvláštnosti pri tvorbe štruktúrnych preskupení chromozómov vedú ich prestávky k zmene pozície génov, čo je sprevádzané zmenou ich expresie (obrázok 22). Dominantný gén A, ktorý sa ukázal byť blízky miestu pretrhnutia chromozómov, vo väčšej miere stráca svoj vplyv ako gén B, ktorý sa nachádza ďalej od miesta prasknutia. Inými slovami, oslabenie účinku dominantného génu je úmerné vzdialenosti medzi ním a bodom zlomu chromozómu.
V iných prípadoch môže chromozomálna prešmykovanie:
Oddeliť transkripčnú jednotku od regulačnej oblasti, pričom úplne neutralizuje jej účinok na gén (neprítomnosť zosilňovača znižuje alebo eliminuje transkripciu v zodpovedajúcom tkanive); na druhej strane oddelenie génu a jeho tlmiča vedie k abnormálnemu zvýšeniu expresie;
Pohyb gén Enhancer región iný gén spôsobujúci nevhodnou expresiu (napr., Burkittov lymfóm translokácia presunie c-myc génu pod kontrolou imunoglobulínu enhanceru);
Presuňte gén a jeho regulačné sekvencie na iný gén, ktorý znižuje úroveň expresie prvého génu;
Vytvoriť mozaikový efekt pozície (napríklad červená a biela farba očí v Drosophile).
 Obr. 22.Účinok polohy v prípade ruptu chromozómu:
Obr. 22.Účinok polohy v prípade ruptu chromozómu:
C - centromér; p a q - krátke a dlhé ramená chromozómu; X - gén
lokuse; A a B - dominantné gény
Ďalším príkladom prejavu vplyvu situácie - jednotlivcov s krvnými skupinami na systéme Rh. Najmä jedinci s komplexom CDE / cDe génu majú rovnaké gény ako jedinci s komplexom cDe / CDE génu (kde veľkým písmenom je dominantný gén a veľkým písmenom je recesívny gén). Prvé z nich majú veľa antigénu E a malého antigénu C, ktoré majú opačný účinok, čo sa vysvetľuje nerovnakou lokalizáciou génov C a E na tom istom chromozóme alebo na rôznych chromozómoch.
Ďalší príklad - autozomálne dominantné plocha typu myodystrophy lopatochnoplechevaya 1A v dôsledku delécie variabilný oblasti v subtelomeric chromozóme dlho ramenové časti (umiestnenej v 4q35 segmente), ktorý neobsahuje gény, ale má nukleotidovej sekvencie 20-250 KBP veľkosti. Delécia týchto sekvencií vedie k zmene štruktúry heterochromatínu v tejto oblasti chromozómu a sprostredkovanej inhibícii transkripcie blízkeho génu (FRG1) umiestneného proximálne od bodu ruptúry chromozómu.
Zavolajte niekoľko dedičných ochorení súvisiacich s vplyvom situácie. Medzi ne patrí:
Aniridium ako výsledok delécie génu lokalizovaného v lokuse
Holoprocephalus - mutácia v géne SSN (7q36);
Satre-Kotzen craniosynostóza - mutácia v géne TWIST (7p21);
Cephalopolisindactyly Greig v dôsledku mutácie SLIZ
Riegerov syndróm - gén PITX2 (4q25-q27);
XY zvrátenie pohlavia - mutácia v géne SRY (Xq).
Genocopying a jeho príčiny
Rovnaký znak (skupina znakov) je spôsobený rôznymi genetickými príčinami (alebo heterogenitou). Takýto účinok bol na návrh nemeckého genetika H. Naftheima získaný v polovici 40. rokov 20. storočia. meno genokopirovaniya.Sú známe tri skupiny príčin genocídy.
Príčiny prvej skupinykombinuje heterogénnosť kvôli multi-lokalite alebo akcie rôznych génov nachádzajúcich sa v rôznych miestach na rôznych chromozómoch. Napríklad medzi dedičnými metabolickými ochoreniami komplexných cukrov - glukózových aminoglykánov sa rozlišuje 19 typov (subtypov) mukopolysacharidóz. Všetky typy
sú postihnuté poruchami rôznych enzýmov, ale prejavujú sa tým istými (alebo podobnými) symptómami gargoylového dysmorfizmualebo feno-typ zvonenia Quasimodo - protagonista románu "Notre-Dame de Paris" klasikou francúzskej literatúry Victor Hugo. Podobný fenotyp sa často pozoruje u mukolipidózy (poruchy metabolizmu lipidov).
Ďalším príkladom multilokusu je fenylketonúria. Ktorí sú označené nielen svojím klasickým štýlom kvôli nedostatku fenylalanín-4-hydroxylázy (12q24.2), ale tiež tri atypické formy: jedna spôsobená nedostatkom digidropteridinreduktazy (4r15.1) a ďalšie dva - deficit enzýmov piruvoiltetragidropterin syntetázy a tetrahydrobiopterínu (čo zodpovedá gény, ktoré ešte neboli identifikované).
Ďalšie príklady multilokusová: glycogenoses (10 genokopy), Ehlers-Danlosův syndróm (8) neyrofibramatoz Reklingauzena (6), vrodené hypotyreózy (5), hemolytická anémia (5), Alzheimerova choroba (5), Bardet-Biedl (3) syndróm, rakoviny prsníka (2).
Príčiny druhej skupinykombinuje intralokálnu heterogenitu. Je to buď viacnásobná alelizmus (pozri kapitolu 2), a to buď genetické zlúčeninyalebo dvojitých heterozygotov s dvoma identickými patologickými alelami v identických lokusoch homológnych chromozómov. Príkladom druhej - heterozygotná beta-Thalassemia (11r15.5) je tvorená v dôsledku delécie dvoch génov kódujúcich p-globínu reťazca, čo vedie k vyššiemu obsahu hemoglobínu HbA 2, a zvýšila (normálne) hladiny HbF hemoglobín.
Príčiny tretej skupinykombinuje heterogenitu spôsobenú mutáciami v rôznych miestach toho istého génu. Príkladom je cystická fibróza (7q31-q32), ktorá sa vyvíja v dôsledku prítomnosti takmer 1000 bodových mutácií v géne zodpovednom za túto chorobu. S celkovou dĺžkou génu cystickej fibrózy (250 tisíc N. p.) Očakáva sa, že v ňom bude detekované až 5 000 takýchto mutácií. Tento gén kóduje proteín zodpovedný za transmembránový prenos chlórových iónov, čo vedie k zvýšeniu viskozity sekrécie exokrinných žliaz (pot, slinných, sublingválnych atď.) A blokovania ich kanálikov.
Ďalším príkladom je klasická fenylketonúria, v dôsledku prítomnosti 50 bodových mutácií v géne kódujúcom fenylalanín-4-hydroxylázu (12q24.2); Celkovo toto ochorenie má zachytiť viac ako 500 bodových mutácií génu. Väčšina z nich vzniká
kvôli polymorfizmu pozdĺž dĺžky reštrikčného fragmentu (RFLP) alebo počtu tandemových opakovaní (VNTP). Bolo preukázané, že hlavnou mutáciou génu fenylketonúrie v slovanských populáciách je R408 W /
Pleiotropický účinok
Vyššie uvedená nejednoznačnosť povahy vzťahov medzi génmi a znakmi je tiež vyjadrená v pleiotropický účinokalebo pleiotropného účinku, keď jeden gén spôsobuje vytvorenie viacerých znakov.
Napríklad autozomálny recesívny gén telangiektázie ataxie, alebo louis Bar syndróm(11q23.2) je zodpovedný za súčasnú deštrukciu najmenej šiestich telesných systémov (nervový a imunitný systém, koža, dýchacie a gastrointestinálne sliznice a očná spojivka).
Ďalšie príklady: gén bardet-Beadleov syndróm(16q21) spôsobuje demenciu, polydaktívne, obezitu, degeneráciu pigmentu sietnice; Fanconiho anémia gén (20q13.2-13.3), riadenie aktivity topoizomerázy I, čo spôsobuje anémiu, trombocytopénia, leukopénia, mikrocefália, polomer aplázia, hypoplázia záprstia Aj prst malformácií srdca a obličiek, hypospádia, vekové škvrny koža, zvýšená krehkosť chromozómov.
Priraďte primárnu a sekundárnu pleiotropiu. Primárna pleiotropiav dôsledku biochemických mechanizmov účinku mutantného enzýmového proteínu (napríklad nedostatok fenylalanínu-4-hydroxylázy pri fenylketonúrii).
Sekundárna pleiotropiakvôli komplikáciám patologického procesu, ktorý sa vyvinul ako výsledok primárnej pleiotropie. Napríklad vzhľadom na zvýšenú tvorbu krvi a hemosiderózu parenchýmových orgánov vyvíja pacient s talasémiou zhrubnutie kostí lebky a hepatolíniového syndrómu.
Alelelická séria
Bodové mutácie jedného génu môžu spôsobiť vývoj nielen jedného komplexu symptómov, ale aj rôznych symptómov. V druhom prípade zohrávajú úlohu alelické série rovnakého génu.Napríklad mutácie génu adrenoreceptora spojené s chromozómom X spôsobujú kennedyho choroba,ak zachytia oblasť trinukleotidových opakovaní v exóne 1, ale môžu viesť k syn-
morris Dromaalebo feminizácia semenníkov (fenotypicky - dievča, genotypicky - chlapca, to znamená prítomnosť ženského fenotypu so samčím karyotypom), ak sú zachytené iné nukleotidové sekvencie tohto génu (mutácia v doméne viažucej si androgén).
Ďalším príkladom heterogénnosti alelických sérií sú bodové mutácie génu tyrozínkinázového receptora - RET, spôsobujúce štyri dedičné choroby: familiárny medulárny karcinóm štítnej žľazy, Hirschsprungova choroba, mnohočetná endokrinná neopláziadva typy - 2A (MEN2A) a 2B (MEN2V).
Hoci opísaný fenomén bol objavený relatívne nedávno, je už známe viac ako 100 monogénnych ochorení spôsobených mutáciami jediného génu v alelickom sérii. Ich dôvody sú:
Rôzne nukleotidové sekvencie (rôzne body génu), ktoré kontrolujú funkčne odlišné proteínové domény;
Prítomnosť v tej istej mutantnej alele modifikujúcej gén (alelický polymorfizmus);
Vplyv genetického prostredia na manifestáciu mutantnej alely (vrátane interakcie mutantnej alely s alelami jedného alebo viacerých modifikujúcich génov).
Fenokopirovanie
Fenokopirovanie- to je kopírovanie komplexu príznakov nedeštruktívnej choroby (vytvorenej pod vplyvom environmentálnych faktorov) ako symptomatického komplexu dedičnej genetickej choroby.
Napríklad príznaky endemickej bolesti korešpondujú s príznakmi vrodenej hypotyreózy, ale v prvom prípade je dôvodom nedostatok (alebo úplná neprítomnosť) anorganického jódu v pitnej vode a potravine (t.j. je fenokopy), a v druhom - rôzne genetické faktory (m. e) existuje genocída).
Ďalším príkladom duplikácie génov a symptomatológie fenokopy je slepota. Je známe, že vízia je riadená skupinou génov (génovej siete), ktoré navzájom ovplyvňujú počas ontogenézy a zabezpečujú vývoj a udržiavanie funkcií očí a mozgu. Rôzne dôvody porušenia integrity tohto systému môžu spôsobiť rovnaký výsledok (slepota).
Takže v dôsledku genocídy sa môže vyvinúť slepota v dôsledku defektov šošoviek určených určitými génmi, defektov sietnice definovaných inými génmi a defektov rohovky určených tretími génmi.
V dôsledku fenokópií sa v dôsledku exogénneho vplyvu životného prostredia môže vyvinúť slepota z dôvodu nedodržiavania noriem osvetlenia obytných a priemyselných priestorov alebo následkom poškodenia očí.